 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
项目文章|AG代理基因nanopore全长转录组测序助力发表植物学顶级期刊the plant cell
2022年8月3日,中科院植物所林荣呈研究组以“The RNA helicase UAP56 and the E3 ubiquitin ligase COP1 coordinately regulate alternative splicing to repress photomorphogenesis in Arabidopsis”为题的研究论文在期刊“The Plant Cell”上在线发表,该研究阐明了UAP56与COP1共同通过调节下游基因可变剪接来调控光形态建成的分子机制和调控网络,增加了对植物光信号转导机制的认识。AG代理基因在本研究中承担了全长转录组测序及分析工作。

研究背景
选择性剪接(AS)是一个关键的转录后过程,可调控基因转录成不同的蛋白质变体,多用于蛋白质组多样性和基因调控研究。先前的转录组学分析表明,光在植物中诱导大量AS。然而,只有少数参与调节光响应的剪接调节器被确定。本研究发现RNA解旋酶UAP56在体内外与光信号通路中的核心角色E3泛素连接酶 COP1相互作用,并通过Nanonpore全长转录组测序分析了UAP56和COP1共同调节许多基因的表达水平以及许多常见前信使RNA(mRNA)的剪接,随后又证明了UAP56和COP1与常见的小核RNA和靶mRNA结合。
材料与方法
黑暗5天和红光1小时的col-0, UAP56 RNAi d3#2和cop1-4幼苗,每个基因型分别收集3个独立样本,进行Nanonpore全长转录组测序和相关分子实验。
研究结果
UAP56对拟南芥的生长发育至关重要
通过RNAi和CRISPR/Cas9的方法分别获得UAP56的突变体株系,并通过免疫印迹分析其表达量下降(图1A)。随后使用其来研究UAP56的生物学功能。与Col-0相比,所有uap56株系显示出与野生型相同的开花时间,但显示出发育缺陷(图1 F-G)。
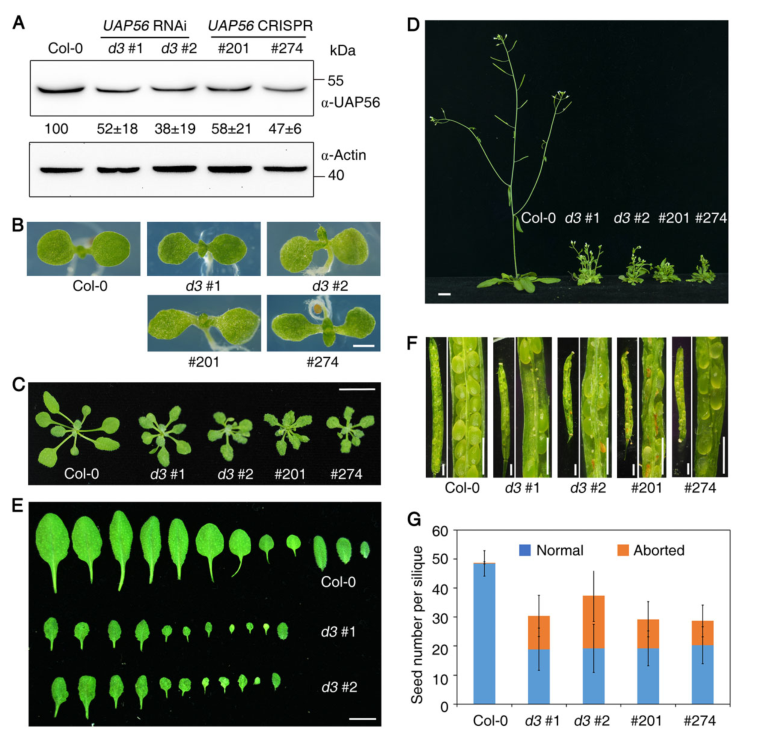
图1 UAP56基因敲除导致多效性发育缺陷
UAP56抑制光形态发生
当在连续红光、远红光、蓝光和黑暗条件下生长时,uap56转基因株系显示出比Col-0更短的下胚轴,并在黑暗中无法形成顶端钩。而35Sp:UAP56a GFP幼苗与Col-0相同(图2 A-B)。uap56转基因株系在连续远红光下生长时积累了更多的花青素,在持续红光照(cR)下生长时叶绿素含量高于Col-0(图2 C-D)。通过RT-qPCR检测了几个关键的光调节基因,结果表明,UAP56负调控光形态发生(图2 E-F)。
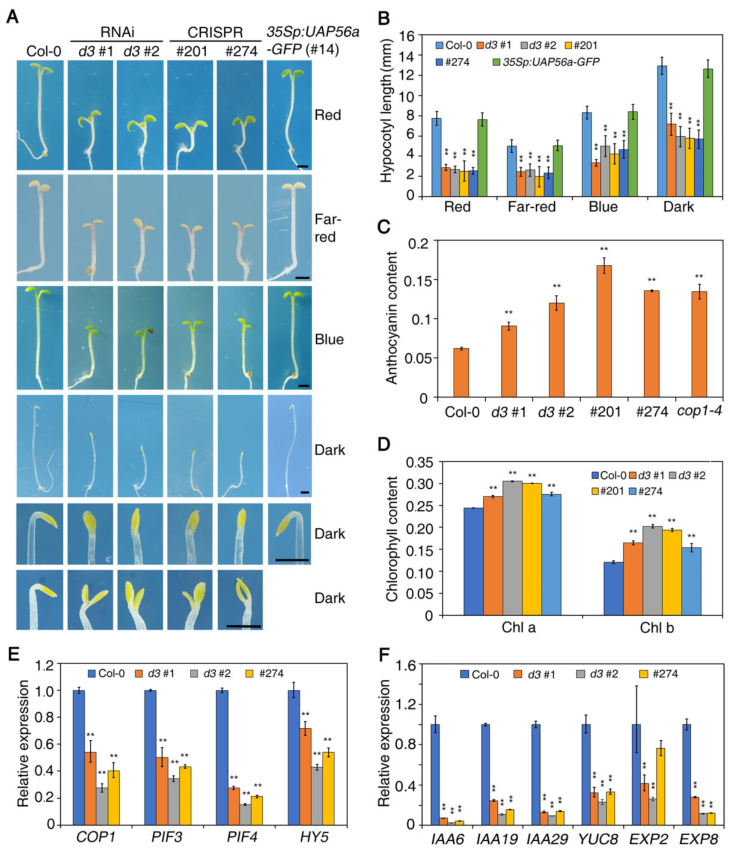
图2 uap56株系的光致形态响应
UAP56与COP1相互作用
COP1是光形态发生的核心抑制因子。通过酵母双杂、BiFC、LUC互补成像(LCI)、co-IP实验和Pull-down实验证明UAP56在体内外都与COP1互作(图3 A-D)。将UAP56 RNAi系与转基因cop1-4进行杂交,其结果表明UAP56和COP1以相加或部分独立的方式调节光形态建成(图3 F-G)。
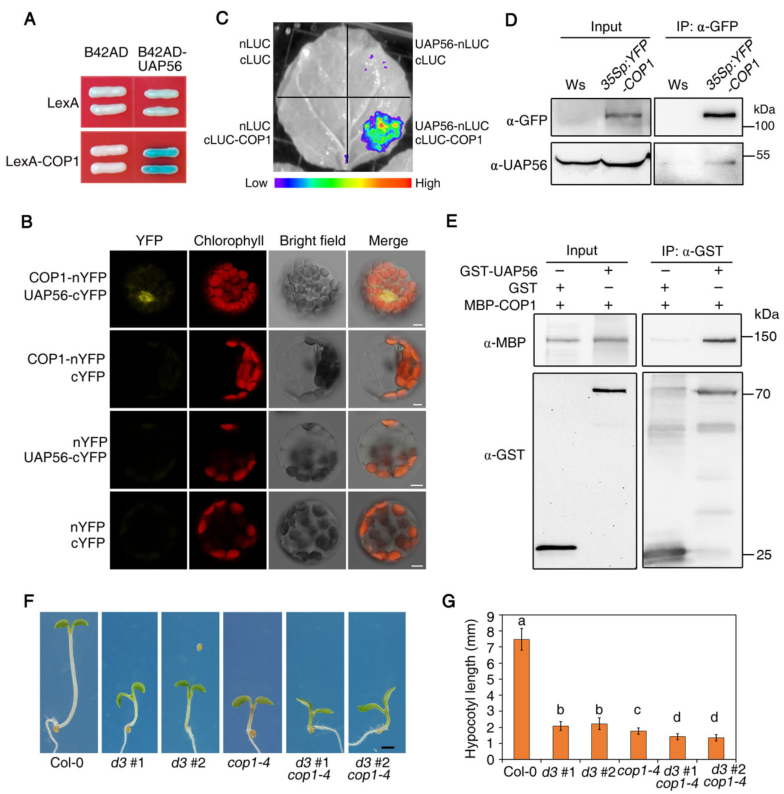
图3 UAP56与COP1相互作用
UAP56和COP1共同调节基因表达
为了研究UAP56和COP1如何共同调控光形态发生,对d3#2和cop1-4以及Col-0进行了Nanonpore全长转录组测序分析。在黑暗和红光照射1小时后,与Col-0相比,在d3#2系中发现了也受COP1的调控基因(图4 A-B)。GO分析显示,UAP56和COP1共同调节与光合作用和光相关的基因表达,以及黑暗或光明中的激素反应(图4 C-D)。随后分析了由UAP56和COP1共同调节的DEGs并选择了6个代表基因通过RT-qPCR来验证Nanonpore全长转录组测序数据(图4 E-H)。
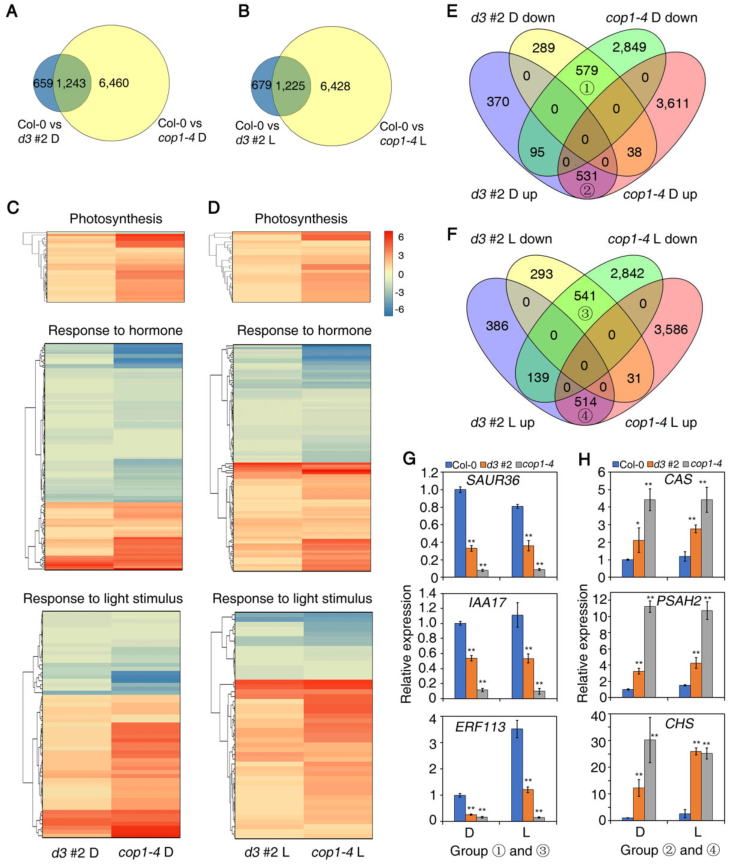
图4由UAP56和COP1共同调节的RNA-seq
UAP56和COP1控制mRNA的选择性剪接
我们通过分析RNA-seq数据证明了UAP56和COP1在拟南芥中也调节AS(图5A)。AS基因的热图表明,d3#2和cop1-4突变体中的剪接模式在黑暗和光明中与Col-0相比有很大不同(图5B)。维恩图显示了414和503个AS事件分别在黑暗和红光照射中由UAP56与COP1共同控制(图5 C-D)。热图表示共调节AS事件的基因在uap56和cop1系中部分重叠(图5 E-F)。
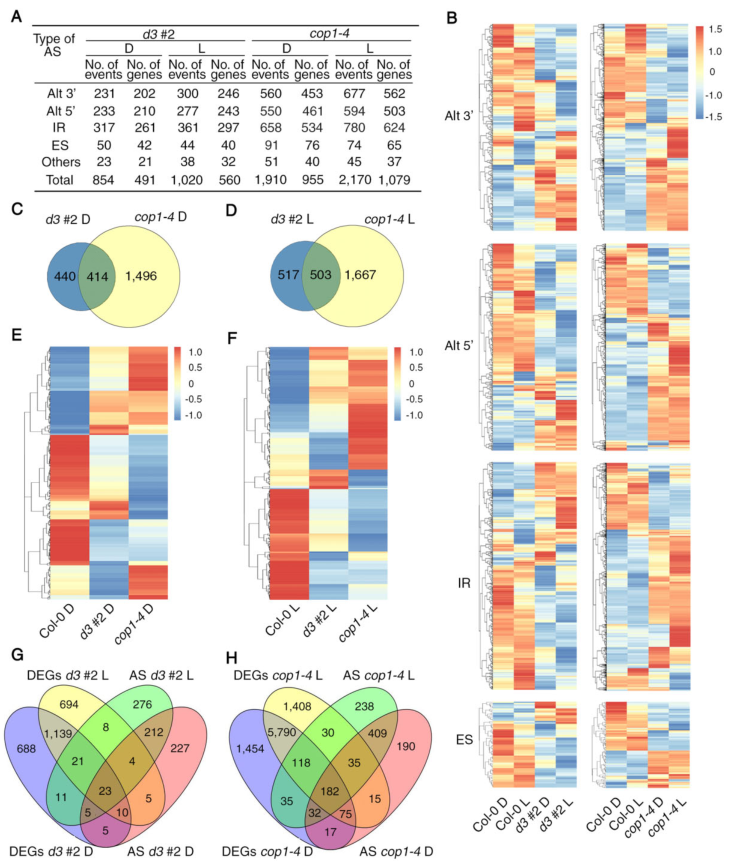
图5由UAP56和COP1控制的AS事件的全基因组分析
UAP56和COP1调节光介导的AS
比较d3#2和cop1-4中受光-暗调节的AS事件,发现433和599 AS事件分别受到黑暗和光明中UAP56突变的特异性影响(图6A)。在cop1中,676和936 AS事件分别在黑暗和光明中发生了特异性改变(图6B)。随后通过散点图表明了 d3#2和cop1-4背景中剪接效率的变化与Col-0有很大不同(图6 C–G)。
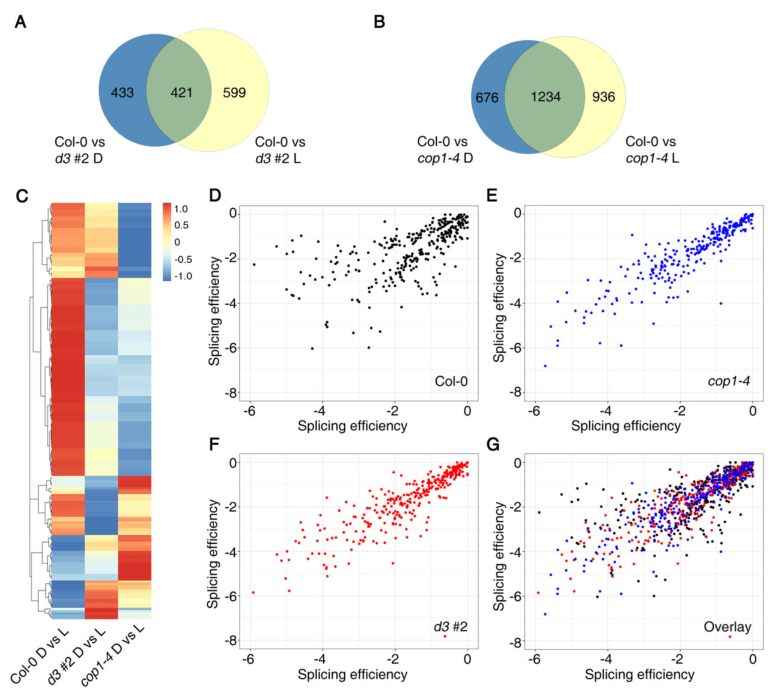
图6 UAP56和COP1在光介导的AS调节中的作用
AS的实验验证
为了验证分析检测到的AS模式,我们对不同光照下生长的Col-0、d3#2和cop1-4幼苗进行了RT-PCR分析。与黑暗相比,cR下d3#2和cop1-4幼苗中RS31、TFIIIA、SR30和DRM1的变化不太明显(图7)。
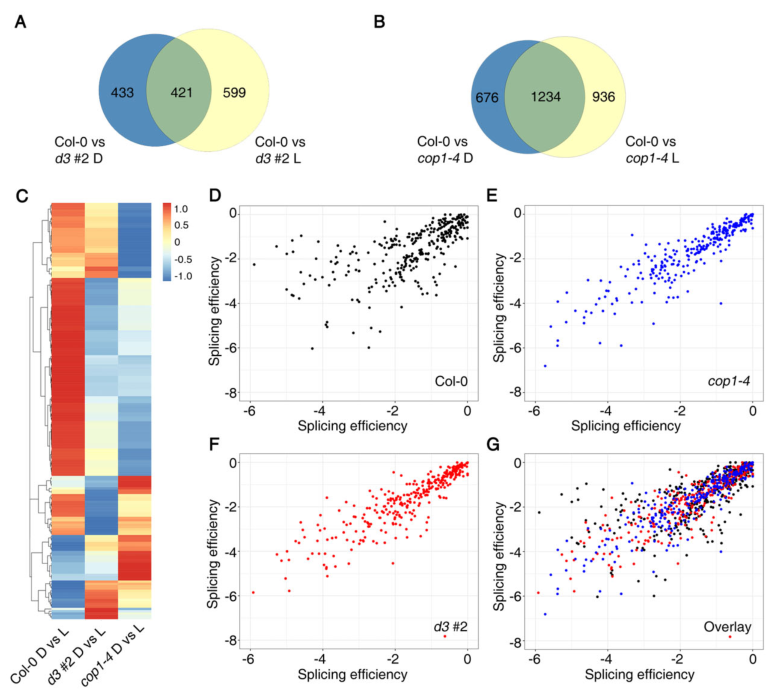
图7通过RT-PCR验证mRNA剪接模式
UAP56与U2AF65相互作用
U2AF是哺乳动物中由U2AF65和U2AF35亚基组成的非snRNP成分,人类UAP56与U2AF67相互作用。因此,我们测试了在拟南芥中UAP56是否与U2AF65相互作用。酵母双杂交、LCI、Co-IP分析表明,UAP56在体外和体内与U2AF65相互作用(图8)。
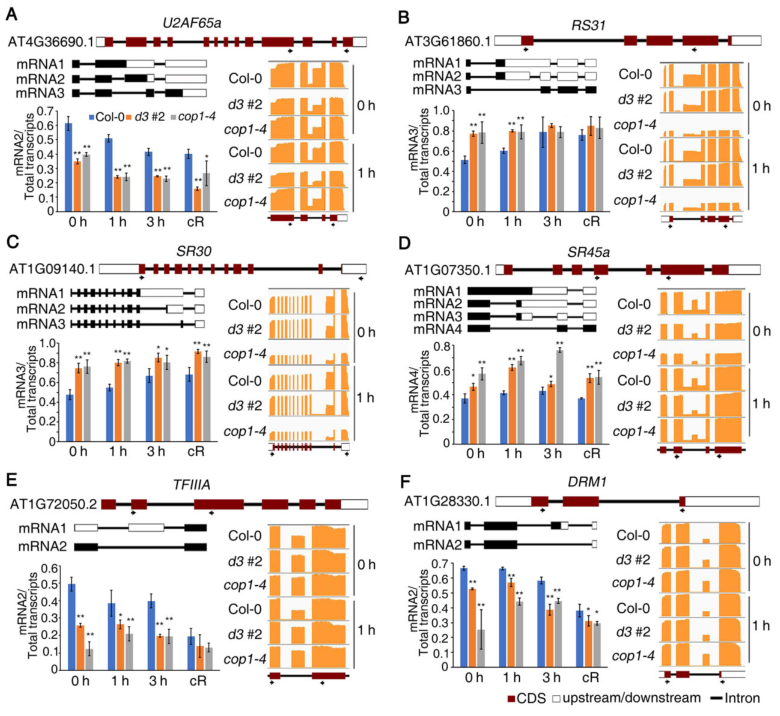
图8 UAP56与U2AF65相互作用
UAP56和COP1与常见snRNA和mRNA结合
我们使用抗UAP56抗体对Col-0种子进行RNA免疫沉淀(RIP)试验,以检测UAP56是否直接与体内的小RNA结合。在黑暗和红光条件下,抗UAP56抗体免疫沉淀了大量的U1、U4、U5和U6 snRNA(图9 A-B)。同样,我们在35Sp:YFP-COP1免疫沉淀物中检测到U1、U2、U4和U5的强烈富集(图9 C-D)。此外,我们比较了缔合强度,发现光照下UAP56富集的U1、U4和U5少于黑暗下的,而COP1在光照下富集的U1、U2、U4、U5和U6少于黑暗下的(图9 F-G)。
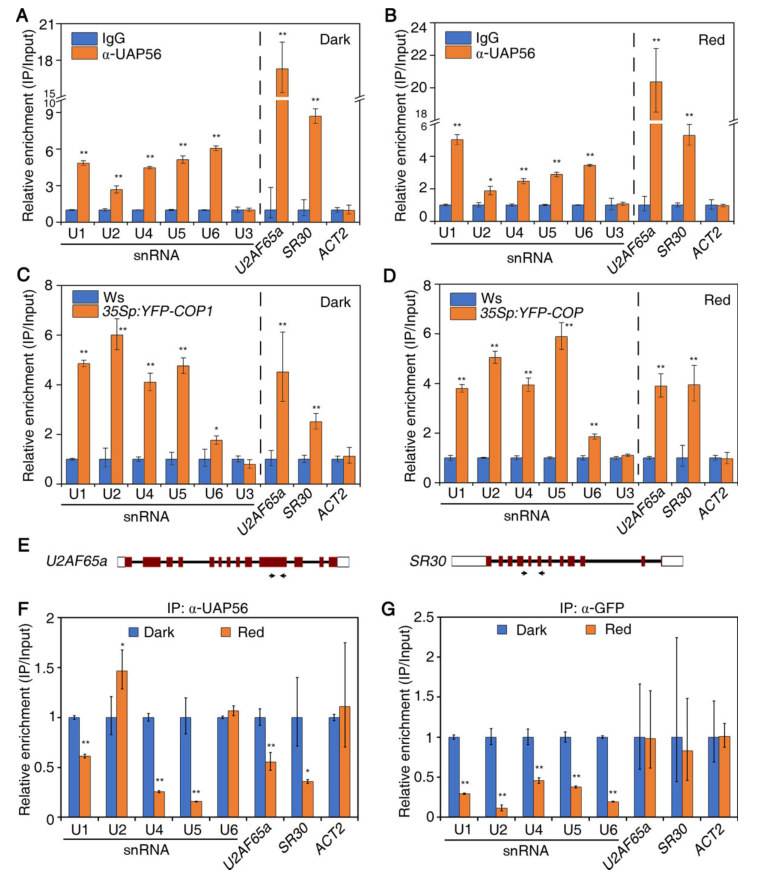
图9 UAP56和COP1在体内结合snRNA和靶mRNA
研究结论
本文证明了UAP56在拟南芥中负调控光发生。UAP56与COP1在体内外均有相互作用。Nanopore转录组测序分析显示,UAP56和COP1共同调节许多基因的表达水平以及许多常见的pri mRNA的剪接。RNA免疫沉淀实验证实UAP56和COP1结合常见的小RNA和靶mRNA。该研究确定了UAP56和COP1在调节植物前体mRNA的AS中的作用和分子机制。


