 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
Plant Cell文献解读 | 稻瘟病菌侵染植物的转录组景观揭示了时间共调控和结构保守的效应因子家族
英文标题:The transcriptional landscape of plant infection by therice blast fungus Magnaporthe oryzae reveals distinct families of temporally co-regulated and structurally conserved effectors
发表期刊: Plant Cell
发表时间:2023.2.18
IF:11.6
作者单位:英国塞恩斯伯里实验室
通讯作者:Nicholas J Talbot
背景介绍
稻瘟病是当前制约水稻生产的重要因素之一,稻瘟病菌(M. oryzae)已进化出具有突破水稻叶片的外角质层,并侵入水稻活体组织的能力,它经历一系列形态转变,在叶片表面分化成侵染细胞——附着胞,随后其侵入性菌丝,渗透进水稻细胞并增殖。效应因子是病原菌分泌的一类蛋白质,可以与植物免疫系统的蛋白相互作用,以抑制宿主的防御系统并使病原菌增殖。
本研究构建了稻瘟病感染的转录调控网络,揭示了植物从最初接种病原菌到疾病症状发展的过程中病原基因的表达变化,并获得了真菌效应因子的完整库。
实验设计
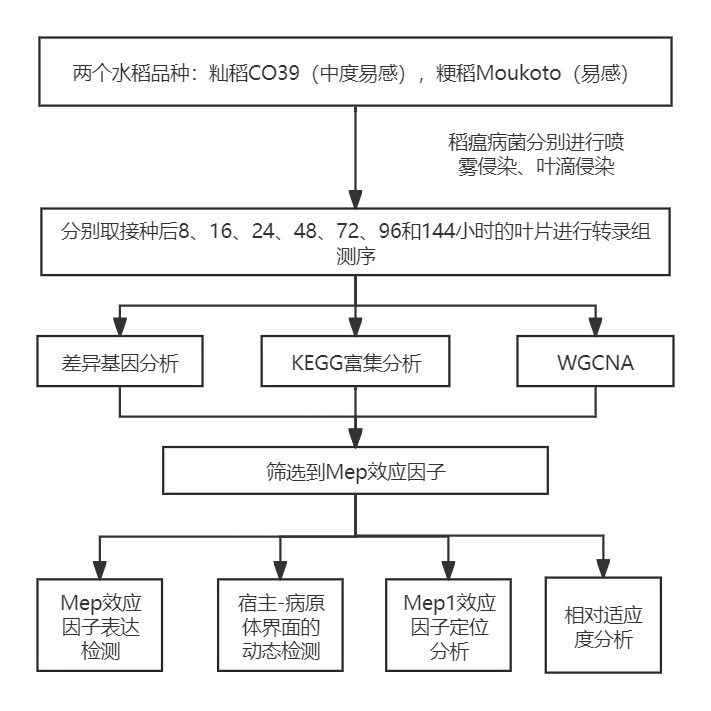
研究结果
1、稻瘟病菌侵染水稻的表型鉴定
通过组织染色实验可以看出稻瘟病菌随着时间推移,侵染规模急剧增加(图1A-B)。对不同感染时期的叶片进行转录组测序,并对水稻和稻瘟病菌的reads进行分类,发现相比喷雾接种组,在叶滴侵染组中真菌转录本比例更高,因此认为叶滴侵染更能代表感染状态(图1C)。
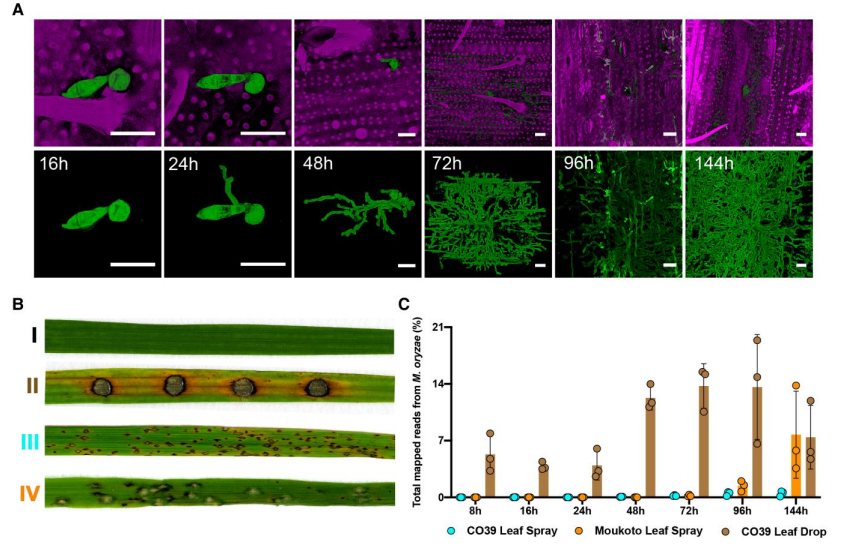
图1.用两种方式接种稻瘟病菌侵染两种水稻过程的转录表达谱分析。
2、稻瘟病期间基因表达的时间动态以及特定的生理转变
利用WGCNA分析了不同感染时间的基因表达,将不同表达模式的基因分为10个模块(图2A)。M1(模块1)主要包含参与真菌生长、分生和孢子萌发的基因,这些基因在孢子时期达到表达峰值;M2主要是与附着胞介导感染相关的基因,侵染8h后表达最高;M3、M4和M5基因分别在16、24和48h达到峰值,主要是与病原菌定殖、发育和功能相关的基因;M6中的部分上调基因与侵染过程中真菌生物量增加相关。M7包含附着孢介导渗透所需的转录因子基因(图2A-B)。不同模块基因的表达模式与其在侵染过程中发挥的功能是一致的。
对不同模块的差异基因进行富集分析,发现在侵入性生长过程中,差异基因富集通路存在从三羧酸循环(M1)到蔗糖代谢(M2, M6,和M9)和磷酸戊糖途径(M8)的转换;M2和M3富集到细胞分裂相关的通路,M4富集到蛋白质加工和输出通路,M5富集到核糖体生物发生通路,这些生物营养通路的富集也表明真菌发生了快速增殖。M6模块显示次生代谢相关基因和细胞壁降解酶的过度表达,与72小时后出现的疾病症状和坏死有关;M7、M8、M9富集到疾病症状发展和发病过程次生代谢通路(图2C)。这些结果表明,基因表达的主要调控变化与初级和次级代谢的开关有关,主要发生在植物感染和生物营养增生到坏死性生长的转变过程中。
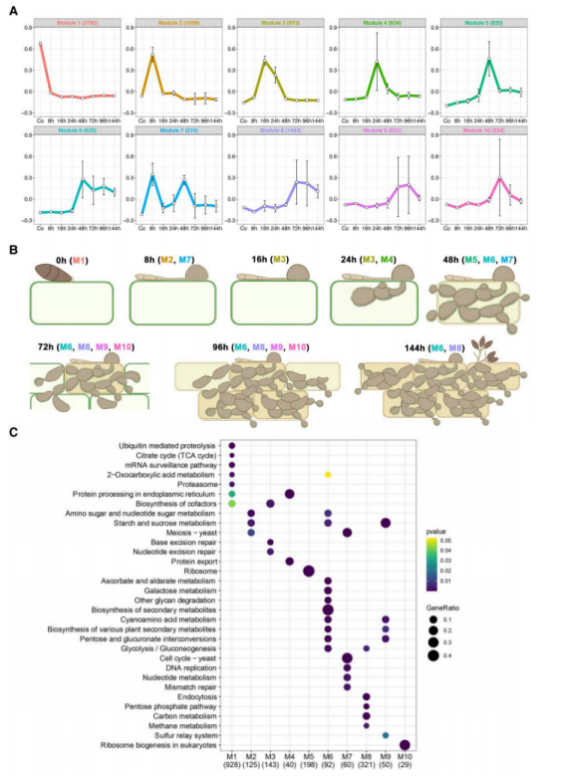
图2.时间共表达分析揭示稻瘟病菌侵染过程中病原基因表达的10个模块。
3、在侵染水稻过程中,稻瘟病菌效应基因家族存在差异表达
利用SignalP从每种接种方法和菌株-栽培互作的数据集中预测分泌蛋白。从喷雾接种的CO39、喷雾接种的Moukoto和叶滴感染的CO39的数据中分别分别鉴定到68、467和847个候选效应基因(图3)。将这些基因分类到不同模块中以揭示这些基因的表达模式(图3A),结果显示在基因表达的初始阶段(M1-M5),不同模块的基因表达模式不同,其中M4和M5中候选效应因子存在过表达。
本研究定义了863个在植物侵染过程中差异表达的Magnaporthe效应蛋白(Mep)基因,进一步使用EffectorP和DeepRedEff算法分别预测到684个和849个效应蛋白,共有546个宿主诱导的Mep基因至少被一种算法预测为效应蛋白(图3C);863个Mep基因在染色体上分布不均(图3D),发现BAS4、Mep4和Mep7等位于端粒附近。
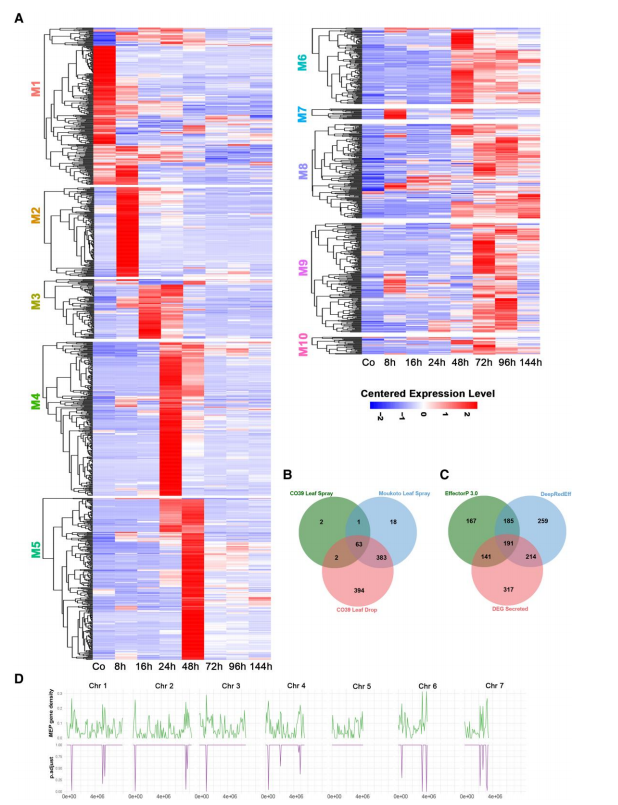
图3.侵染水稻过程中稻瘟病菌分泌蛋白的特异性时间表达。
4、侵染过程中结构保守的M. oryzae效应因子受时间的共调控
根据已知M. oryzae分类结构簇,将863个MEP蛋白分成366个结构簇,选择10个结构最保守的簇,分析它们的编码基因在每个WGCNA模块中的分布,大部分结构保守蛋白在侵染过程中表达。然而MAX-effector簇(簇8)和结构蛋白(簇7)(预测的效应蛋白)在M4、M5中过表达;此外与坏死相关的水解酶(簇1)、葡萄糖苷酶(簇2)和糖基水解酶(簇4和簇6)在M8和M9中过表达。综上,结构保守的候选效应因子在植物侵染期间存在时间共表达(图4)。
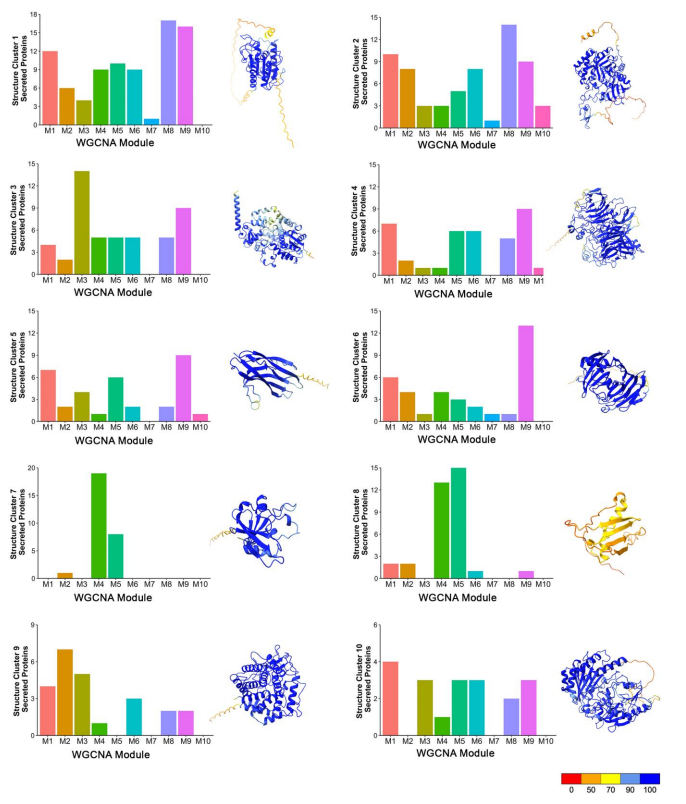
图4.结构保守的M.oryzae效应因子在生物营养侵入性生长过程中时间共表达。
5、M. oryzae的Mep效应因子的阶段特异性表达
随机选择CO39叶滴实验中鉴定的32个Mep基因进行活细胞成像和靶向基因替代实验。研究发现Mep1在植物细胞表面的附着胞孔中形成一个环状构象(图5)。附着胞孔由septin GTPases结合成像,标志着侵染钉出现和极化胞吐的穿透点,故Mep1很可能是在侵染钉穿透角质层时分泌的。角质层破裂后,Mep1-GFP最初聚集在初生侵入菌丝的尖端,随后在侵入菌丝的轮廓处聚集,表明它定位于真菌细胞壁和植物质膜之间的外质体,称为外侵菌丝膜(EIHM)(图5B)。相比之下,M5候选效应物Mep3在感染期间只定位于生物营养界面复合物(BIC),GFP信号在48小时达到峰值(图5C)。此外,还构建了由每个Mep启动子驱动的表达自由GFP的基因融合体,结果表明Mep表达的细胞生物学可视化与转录组分析的预测一致(图5D-E)。
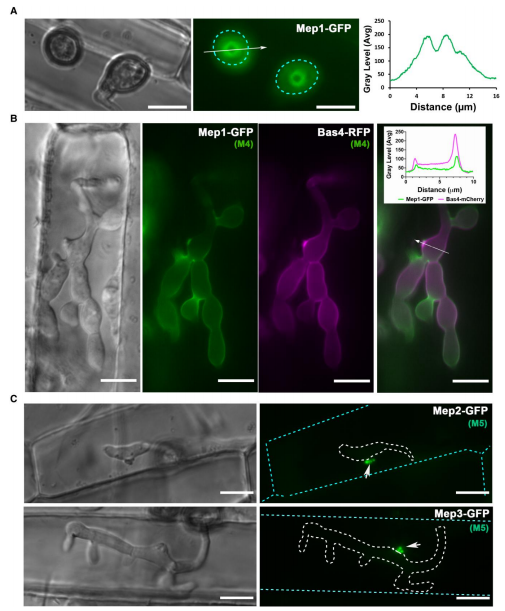
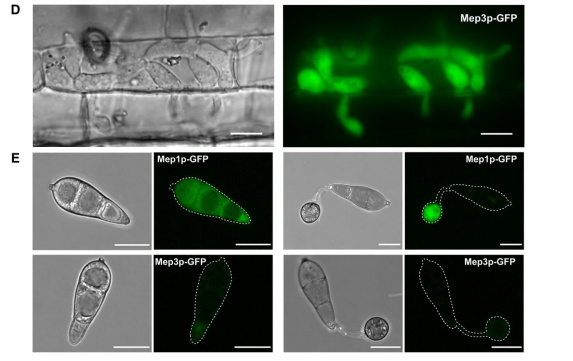
图5.植物受侵染过程中Mep候选物的活细胞成像揭示了附着胞渗透和侵袭生长过程中效应因子的空间定位。
6、植物受侵染过程中宿主-病原体界面的动态变化
为研究植物侵染过程中宿主-病原体界面的动态,用表达Mep1-mCherry的gu11菌株感染表达质膜标记Lti6b-GFP的转基因水稻株系。Mep1-mCherry荧光显示,在以EIHM为界的初始定殖表皮细胞中,存在侵入性菌丝,呈现Lti6b-GFP荧光,这与效应因子传递到真菌细胞壁和EIHM之间的外质体一致。EIHM在植物感染的早期阶段保持完整性(图6A),而一旦真菌入侵邻近细胞,膜完整性就会丧失,导致宿主细胞活力丧失(图6B)。
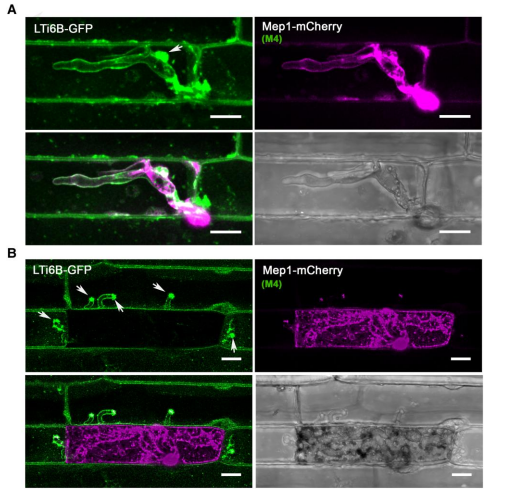
图6. 在植物感染过程中,水稻质膜内陷并积聚在BIC处。激光共聚焦显微照片显示,表达Mep1-mCherry的M.oryzae在表达质膜定位的LTi6B-GFP转基因水稻的表皮叶细胞中定殖。
7、Mep效应因子分泌到BIC被递送到宿主细胞
为研究Mep1效应因子是否可以转移到寄主细胞中,在M.oryzae中表达了荧光效应器融合蛋白和人工添加的C-末端核定位信号(NLS),Mep1-GPF-NLS,并用这种真菌感染了水稻幼苗。尽管该效应因子具有信号肽,通常定位于附着胞孔(图7A),但荧光主要在真菌核中观察到(图7B)。当BIC定位的效应因子Mep3与NLS融合时,在被侵入细胞和邻近细胞的细胞核中观察到Mep3-GPF-NLS信号(图7C)。使用该方法评估的所有BIC定位效应因子都显示易位到宿主细胞,而外质体效应因子,如Mep1,没有易位。当感染了表达Mep-GFP的M.oryzae 的水稻组织暴露于布雷非德菌素A(BFA)时(图7D-E),Mep1-GFP的分泌受到显着抑制,荧光在侵入性菌丝内积聚,BIC相关效应因子(例如Mep2和Mep3)对BFA不敏感,Mep2-GFP信号仍留在BIC中。表明Mep效应因子通过2种不同的分泌途径分泌,具体取决于它们的位置。
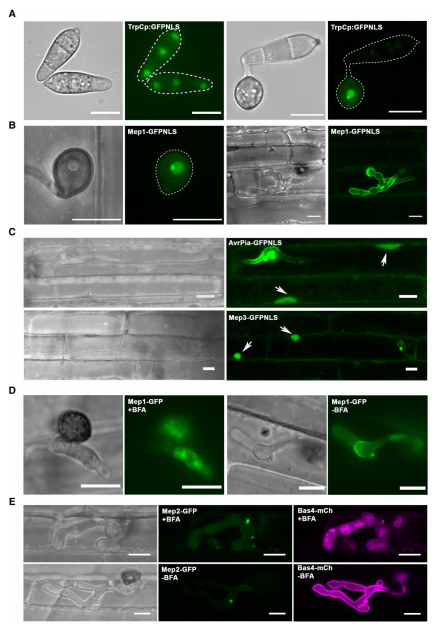
图7. 在BIC处积累的外质体Mep效应因子被转移到宿主细胞中。
8、Mep效应因子有助于病原体适应度
采用相对适应度分析评估效应因子对真菌毒力的贡献。利用组成型启动子TrpC,构建表达细胞质GFP的Guy11菌株。同时,生成第二个由相同TrpC启动子驱动表达的胞质mCherry的Guy11菌株,以及表达由相同TrpC启动子驱动的胞质mCherry的Δmep1突变体。在Guy11菌株的对照实验中,观察到3代后GFP和mCherry分生孢子的回收率没有显著差异(图8b)。相比之下,在Guy11 TrpCp:GFP分生孢子和Δmep1 TrpCp:mCherry分生孢子的混合感染中,Δmep1突变菌株在3轮感染后接近死亡(图8c)。最后,将Mep1基因重新引入Δmep1-MEP1 TrpCp:mCherry菌株,弥补了观察到的适应度缺陷(图8d)。10 d后回收孢子,观察到野生型与突变体保持相同的1:1比例,这表明Δmep1突变体在植物外生长时具有相同的适应度。这些结果表明Mep1参与了M. oryzae引起的稻瘟病,它的缺失对真菌在群体水平上的相对适应度有显著影响。
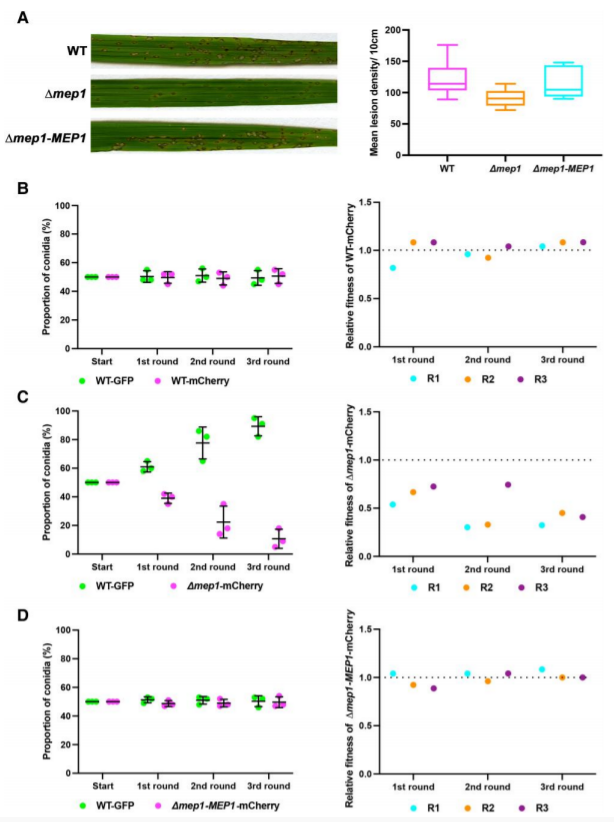
图8. Mep1在植物侵染过程中对病原菌适应性起作用。
结论
基于转录组研究,发现稻瘟病菌具有比以前更广泛的效应因子库,这些效应因子在感染期间的特定时间内表达。这反映在感病的每个阶段,大量与次生代谢、转运蛋白功能和细胞信号传导相关的真菌基因参与表达。同时发现序列无关但结构保守的效应因子在感染期间以协同的方式表达,并发现许多效应因子同时被传递到植物细胞中。此外,本研究开发了一个相对适应度试验,表明了即使单个效应因子也可以增强真菌的侵染毒性。
文章链接:
Yan X, Tang B, Ryder LS, MacLean D, Were VM, Eseola AB, Cruz-Mireles N, Ma W, Foster AJ, Osés-Ruiz M, Talbot NJ. The transcriptional landscape of plant infection by the rice blast fungus Magnaporthe oryzae reveals distinct families of temporally co-regulated and structurally conserved effectors. Plant Cell. 2023 Apr 20;35(5):1360-1385.


