 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
Science Advances详解|深入解析m6A RNA去甲基化酶促进热激下逆转座子转座机制

文章题目:m6A RNA demethylase AtALKBH9B promotes mobilization of a heat-activated long terminal repeat retrotransposon in Arabidopsis
发表期刊:Science Advances
发表时间:2023.11
IF:13.6
通讯作者:Jungnam Cho (中国科学院分子植物科学卓越创新中心)
研究介绍
转座子作为一种DNA遗传元件,广泛分布于真核生物的基因组中,可在染色体中进行位置移动并改变其拷贝数,发生转座现象。转座如若不受控制,往往会影响基因功能发挥,造成基因组的不稳定性。正常情况下,转座子受到DNA甲基化和组蛋白修饰的抑制或沉默,而在环境胁迫下,转座子可被释放和激活,以增加遗传多样性和进化适应性。其中最为典型的便是拟南芥中Ty1/Copia类逆转座子Onsen逆转座子可在热激条件下被转录激活,并调控下游基因响应胁迫。
目前已有的研究大多关注甲基化在转录水平对拟南芥逆转座子Onsen的影响,而逆转座子Onsen对植物生长和发育的转录后修饰及调控的影响仍鲜有报道。近年来,m6A甲基化等转录后修饰成为植物科学领域的研究热点之一。本研究围绕m6A去甲基化酶AtALKBH9B对拟南芥中逆转座子Onsen 的转座调控机制,深入解析了RNA甲基化修饰调控植物响应胁迫过程的内在机理。
研究结果
1.转座子Onsen含有m6A修饰并受m6A修饰调控
大量文献表明,在植物中,尽管热激胁迫可以显著激活转座子的转录,但是很少观察到转座发生。此前在人类有关的研究中发现m6A修饰参与逆转座子调控,由此推测植物中也可能存在类似机制。作者分析现有拟南芥花蕾热激胁迫处理后的m6A RIP-Seq数据,发现热激后m6A修饰在终止密码子附近分布密集,而在转录起始位点则分布较少(图1A),同时鉴定到多个仅在热激胁迫下存在m6A修饰的转录本(图1B),其中转座子占比较高(图1C)。通过对相应样本的转录组数据分析发现,转座子中含有m6A peaks较多的转录本占比更高,包括Onsen逆转座子家族(图1D)。表明,转座子RNA更易受到m6A修饰调控,并且m6A修饰可能参与转座子抑制。
Onsen作为热激胁迫下的逆转座子,可产生染色体外线性DNA(eclDNA)并插入基因组。m6A RIP-Seq数据表明热激下Onsen转录本的m6A修饰水平显著升高(图1E),并且在拟南芥基因组中Onsen家族均显著富集m6A(图1F)。作者对热激胁迫下拟南芥幼苗进行Direct RNA测序(DRS)验证,发现热激下Onsen转录本中确实存在m6A修饰,并通过对特定区域进行RIP-qPCR检测表明热激胁迫影响转座子的m6A修饰水平(图1G)。
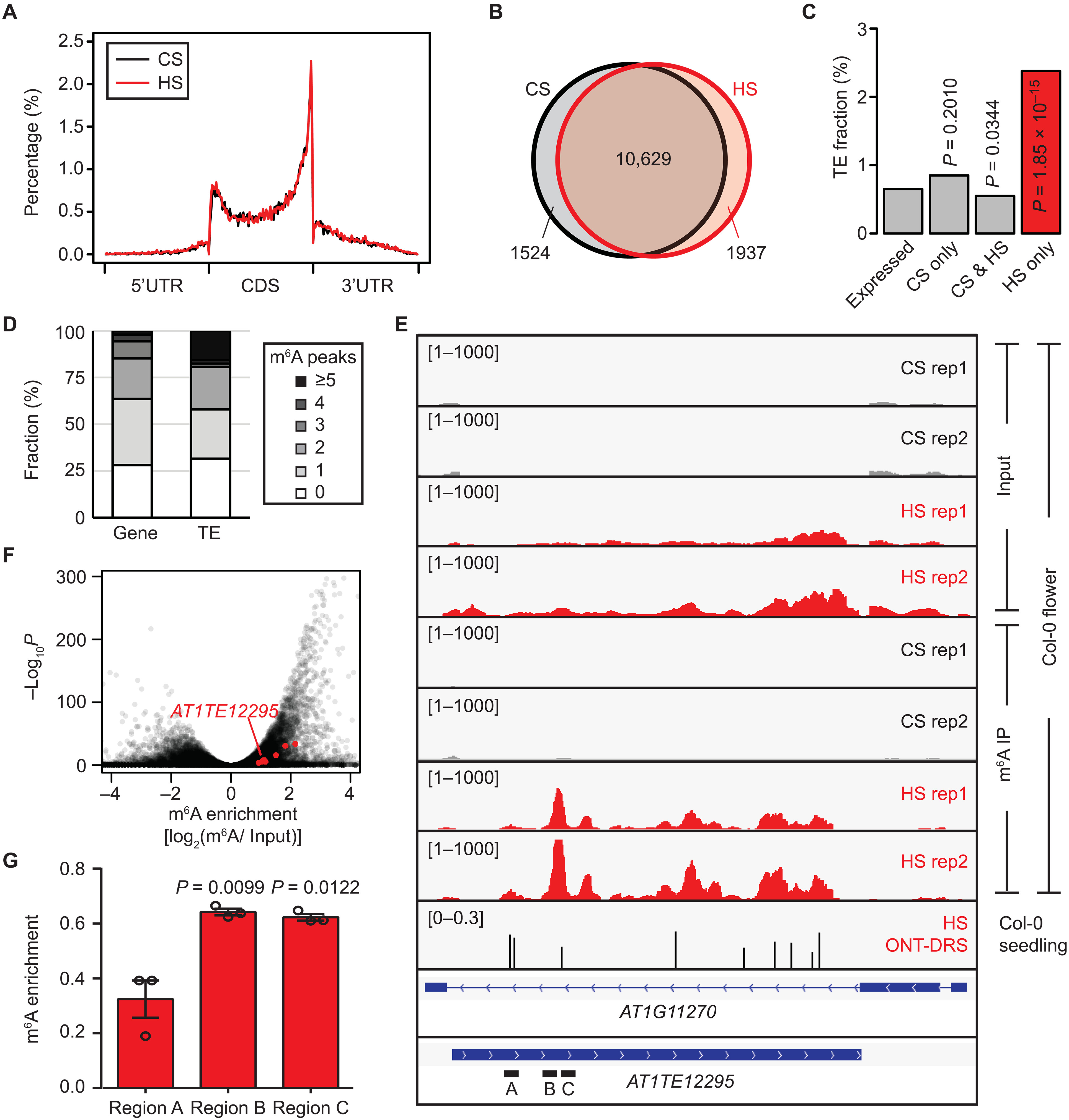
图1:转座子Onsen含有m6A修饰并受其调控
2.m6A RNA去甲基化酶AtALKBH9B参与Onsen转录后调控
作者对现有热激胁迫后拟南芥转录组测序数据分析,推测RNA甲基化可能与热激胁迫响应有关,并从中选择AtALKBH9B基因做进一步的研究。已有研究表明AtALKBH9B参与植物中病毒RNA的去甲基化过程,由于转座子RNA与病毒RNA类似,推测AtALKBH9B也可能调节Onsen RNA。作者分别构建AtALKBH9B插入突变株和缺失突变株(图2A),发现在热激胁迫下,两种突变株的Onsen表达水平均显著上调(图2B),并且在转录组数据和GFP荧光标记实验中得到验证(图2C~D)。随后对热激胁迫下野生型和突变株进行RIP-qPCR检测m6A修饰水平,结果显示突变株中m6A修饰水平显著升高(图2 E~F),由此表明AtALKBH9B可能参与Onsen去甲基化过程。
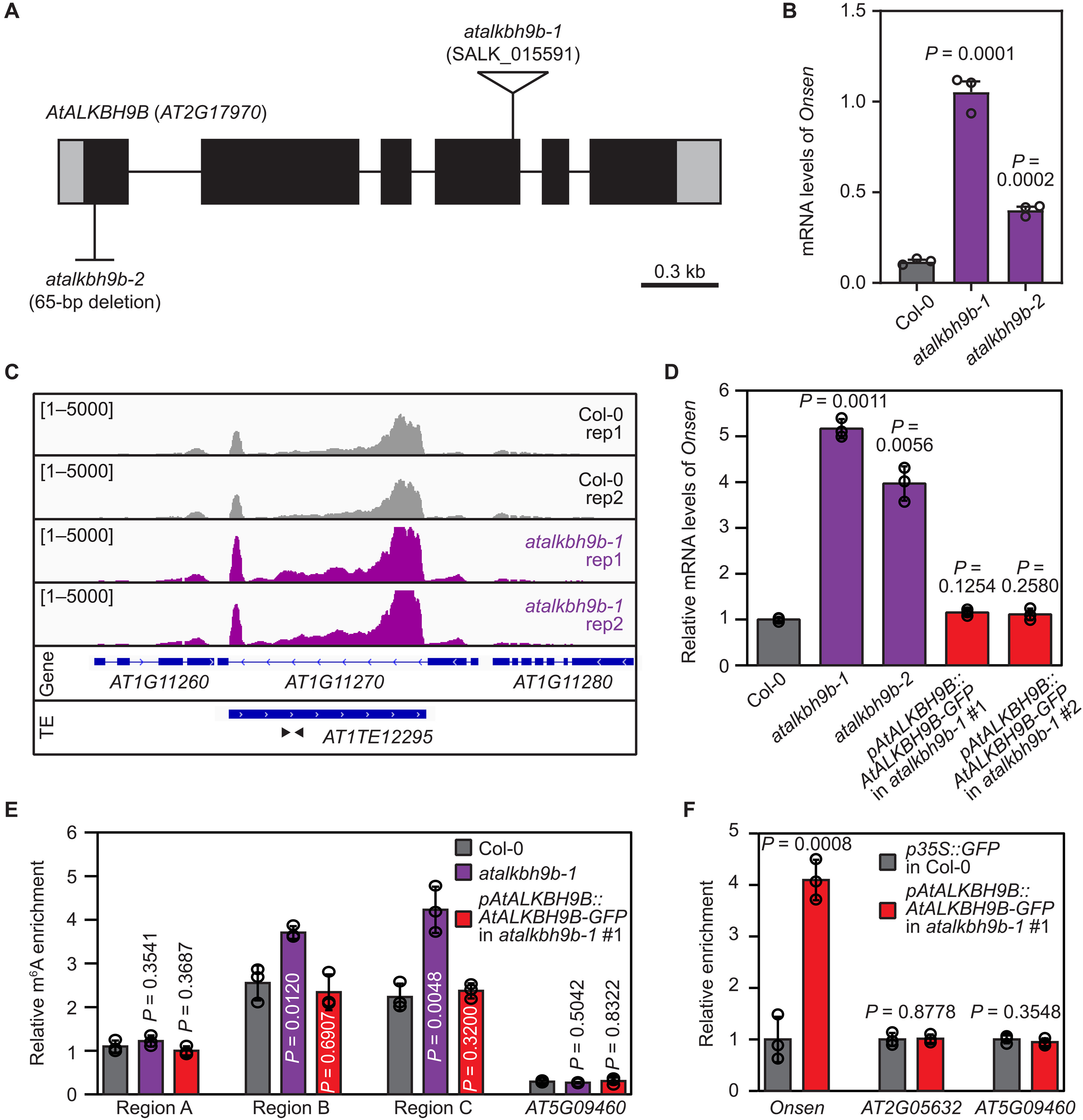
图2:m6A RNA去甲基化酶AtALKBH9B参与Onsen转录后调控
3.AtALKBH9B的缺失影响Onsen转座效率
作者根据AtALKBH9B突变株中Onsen表达上调这一现象推测其转座效率也会有所提升,采用DNA (ALE)–qPCR检测发现,突变株中eclDNA表达水平显著下调(图3A),并且在热激胁迫下的突变株中也表现出表达水平下调(图3B)。进一步通过ddPCR鉴定可知,热激胁迫下突变株及其子代的Onsen拷贝数均显著降低(图3C~D)。以上可以表明AtALKBH9B缺失会影响Onsen转座效率,也由此推测m6A甲基化修饰确实抑制Onsen的转座发生。
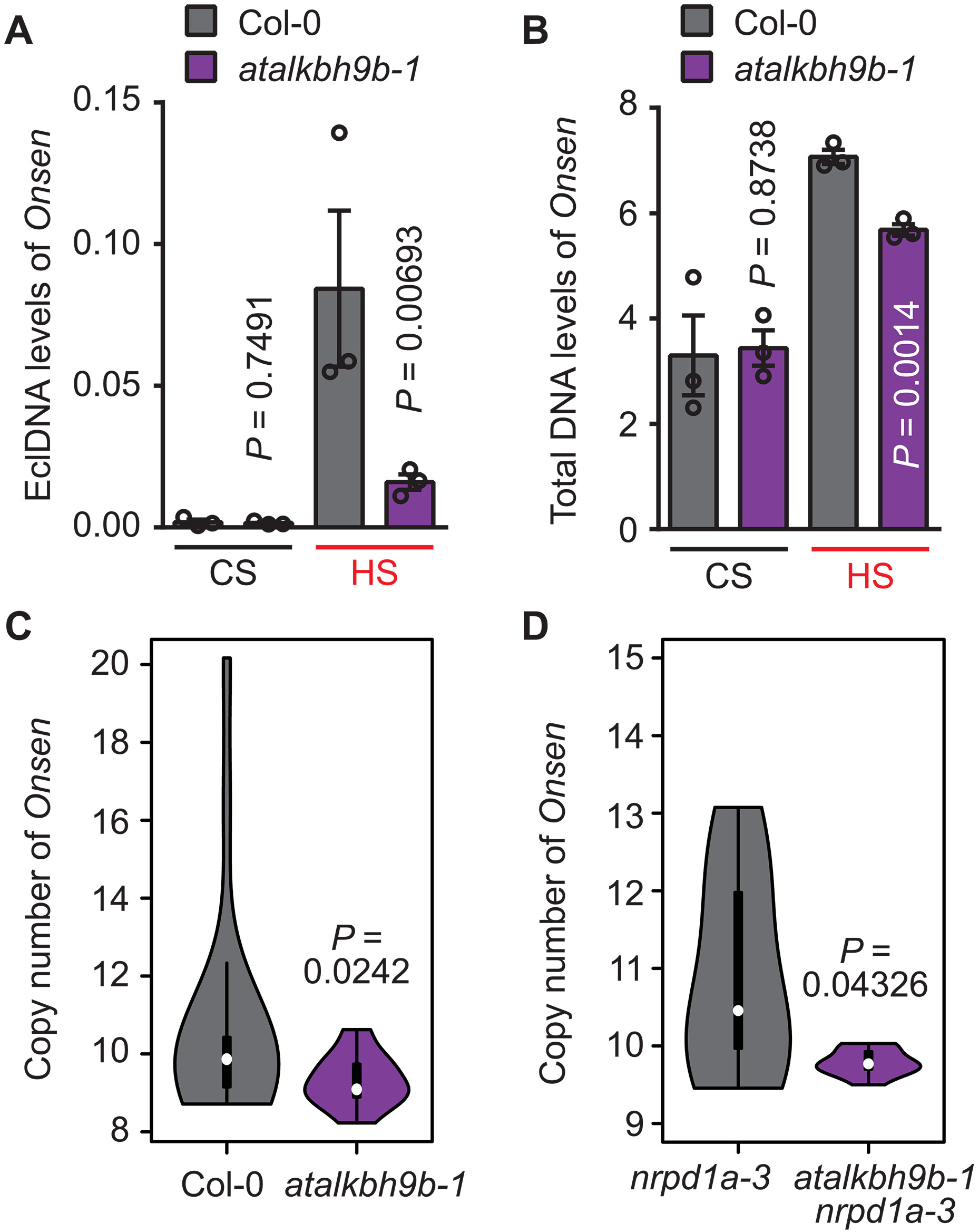
图3:AtALKBH9B的缺失影响Onsen转座效率
4.热激胁迫下AtALKBH9B定位于SG中
应激颗粒(Stress Granules, SGs)是一种动态无膜细胞器,可在应激胁迫下形成并结合RNA和蛋白。此前研究表明在人类中RNA甲基化与SG有关,因此推测AtALKBH9B突变株中高甲基化的Onsen转录本可能定位于SG中。作者将AtALKBH9B与SGS3转入烟草叶片和拟南芥植株中进行共表达定位,发现AtALKBH9B可定位于SG中(图4A~C),并且会与ECT2等多个SG组分发生互作(图4D)。随后作者通过RNA斑点杂交实验和转录组测序尝试确定含有m6A修饰的RNA是否更倾向于定位SG中,发现突变株的SG中转录本的m6A修饰水平更高(图5A~B),并且富集SG的转录本含有更多的m6A peaks(图5C);DRS测序也验证了该结果(图5D)。此外比较突变株中不同转录本的SG富集情况可知,突变株中Onsen 转录本显著富集于SG中(图5E~H)。以上结果间接表明在热激胁迫下,AtALKBH9B突变株中含有m6A甲基化修饰的Onsen转录本可被SG结合,进而影响其转座效率。
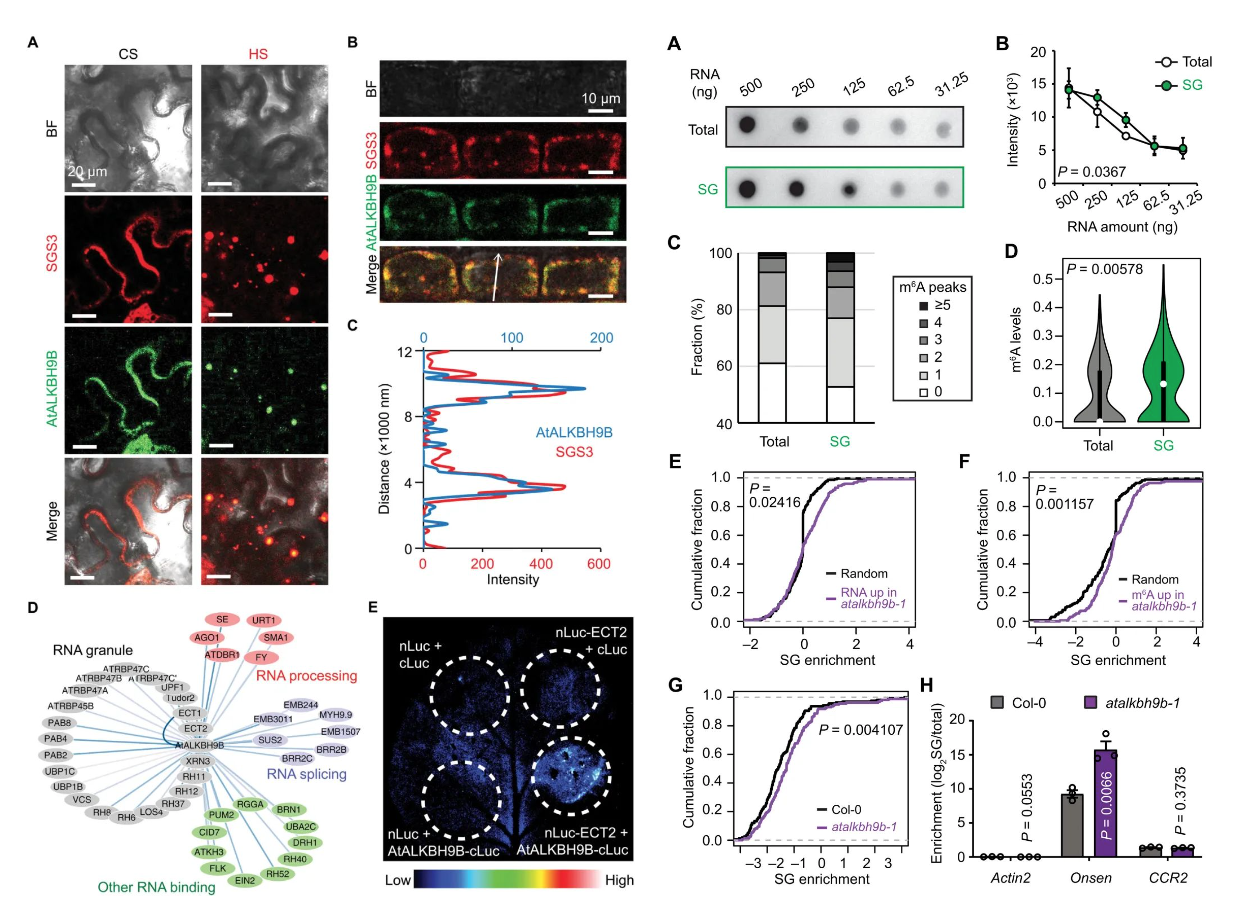
图4~5:热激胁迫下AtALKBH9B定位于SG中(左:图4,右:图5)
5.m6A甲基化抑制VLP组装和eclDNA产生
前述已经明确热激胁迫下含有m6A修饰的Onsen转录本定位于SG中,在去甲基化酶AtALKBH9B的促进下可发生转座。进一步通过荧光偏振和RNA免疫沉淀实验,发现含有m6A 修饰的Onsen转录本与蛋白GAG的结合能力显著降低(图6A)。同时,体外转录以及定量PCR结果显示含有m6A修饰的Onsen RNA的逆转录效率显著降低,影响了cDNA的产生(图6B)。表明,m6A修饰将抑制Onsen RNA参与类病毒颗粒(Virus like particle, VLP)的组装以及eclDNA的产生,最终导致Onsen转座效率降低。
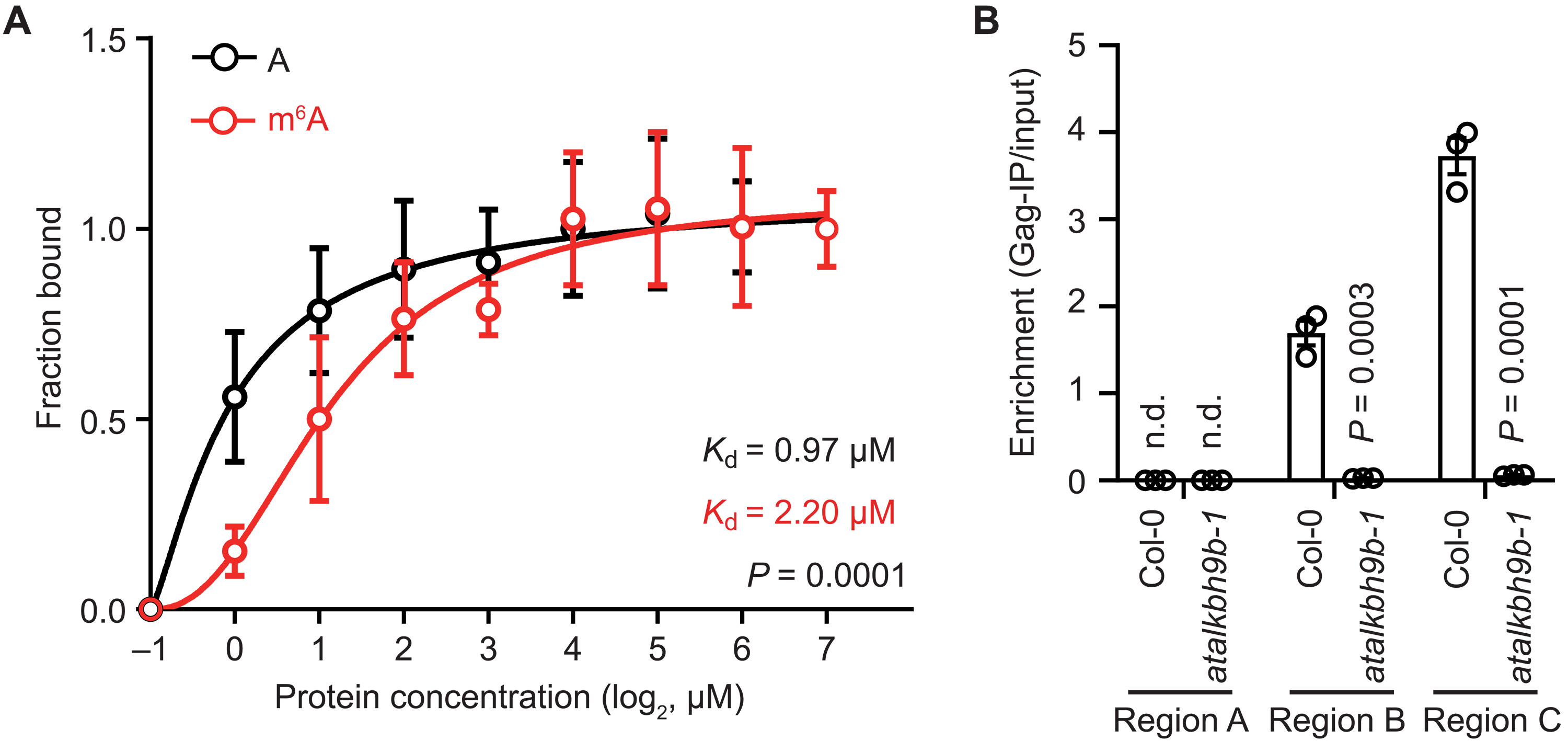
图6:m6A甲基化抑制VLP组装和eclDNA产生
研究总结
本研究结果表明定位于SG的AtALKBH9B可参与调控转座子Onsen RNA去甲基化过程,使其可从SG中释放,完成转座以响应胁迫,进而影响转座发生。总之,本研究揭示了m6A甲基化修饰在抑制转座过程中的作用,并深入解析了转座子如何通过宿主RNA去甲基化酶来对抗m6A介导的抑制机制,为后续转座子及RNA甲基化调控植物响应胁迫的机制研究提供了思路。
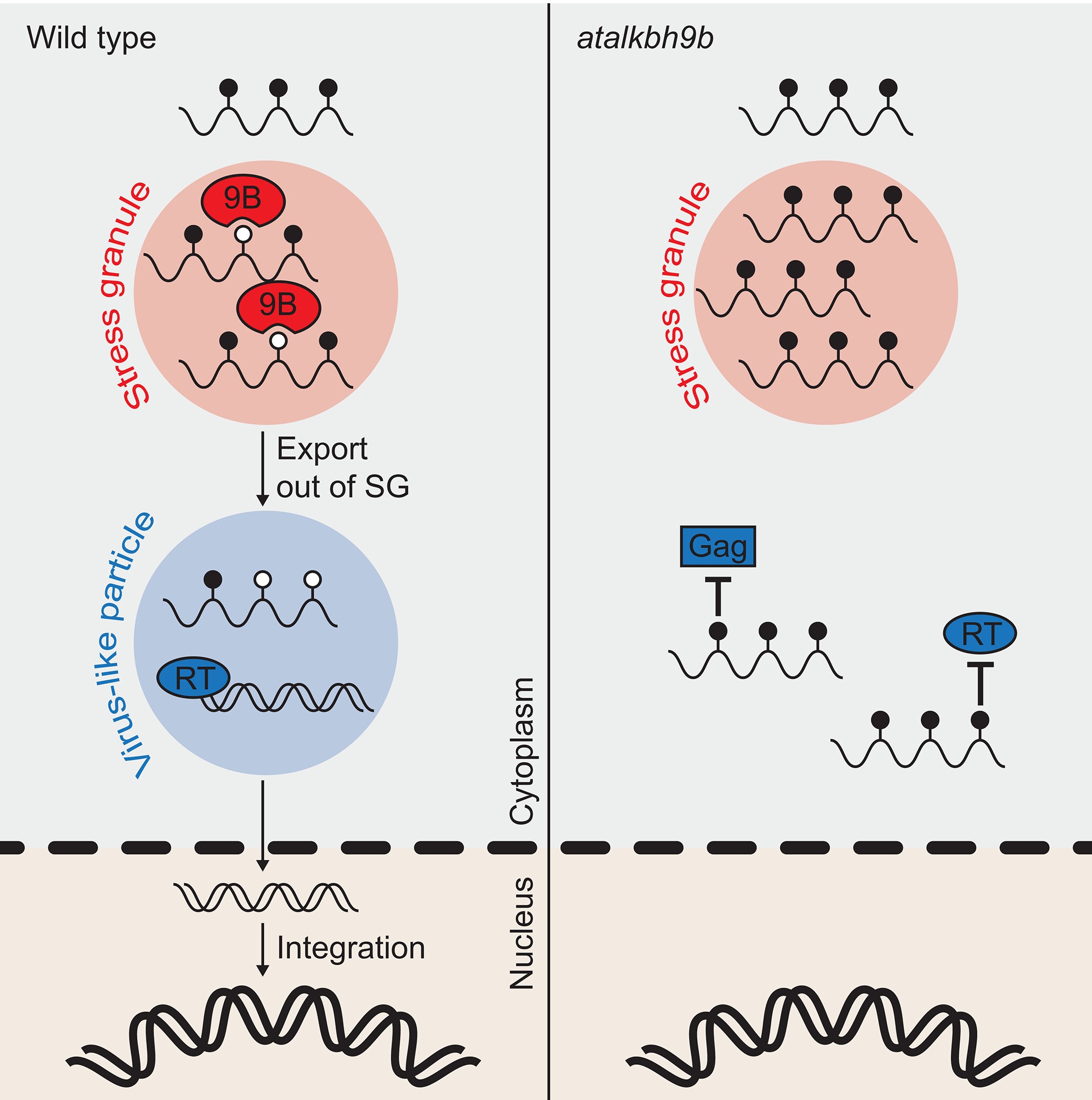
图7:m6A RNA去甲基化酶促进热激下逆转座子转座机制模型
参考文献:
Fan W, Wang L, Lei Z, et al. m6A RNA demethylase AtALKBH9B promotes mobilization of a heat-activated long terminal repeat retrotransposon in Arabidopsis[J]. Science Advances, 2023, 9(48): eadf3292.


