 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
文献解读 | The Plant Cell期刊发表基因组组装助力填补禹氏三角的最后一块:四倍体埃塞俄比亚芥基因组
禹氏三角(英语:triangle of U)是一个关于芸薹属(Brassica)植物的不同物种进化及其相互亲缘关系的理论,是研究芸薹属乃至其它植物多倍化的理想模型。它由6个芸薹属物种构成,包括3个二倍体:白菜(B. rapa, 2n=20, AA)、甘蓝(B. oleracea, 2n=18, CC)和黑芥(B. nigra, 2n=16, BB),以及三者之间相互杂交,经过不断选择进化得到的3个异源四倍体:甘蓝型油菜(B. napus, 2n=38, AACC)、芥菜型油菜(B. juncea, 2n=36, AABB)和埃塞俄比亚芥(B. carinata, 2n=34, BBCC),如图1所示。
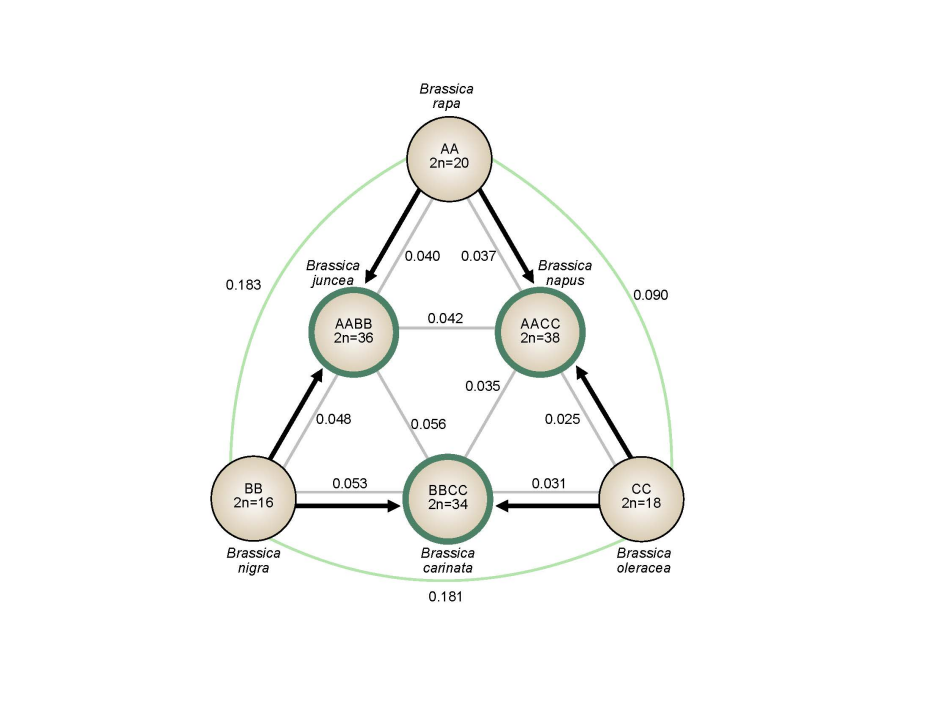
图1 禹氏三角模型下芸苔属植物的遗传关系
埃塞俄比亚芥(Brassica carinata)是一种古老的作物,具有显著的抗胁迫能力和理想的种子脂肪酸结构,可用于生物燃料。该研究报道了B. carinata基因组,填充了禹氏三角模型中的最后一块空缺,确定了农艺性状相关的基因家族,为揭示B. carinata基因组杂交事件,以及研究其优良农艺性状的基因组特征提供了重要的理论依据。

文章题目:The final piece of the Triangle of U: Evolution of the tetraploid Brassica carinata genome
发表期刊:The Plant Cell(IF=12.085)
发表时间:2022.08.12
主要研究结果
1、 高质量B. carinata基因组的组装
该研究利用PacBio、Illumina和Hi-C测序数据组装了B. carinata的基因组,以B. nigra (Bn)和B. oleracea(Co)基因组为参考,分离Bc和Cc的contigs,最终组装得到了1.31 Gb的基因组(图2),挂载到17条染色体上,scaffold N50为78.8 Mb,BUSCO为97.0%。B. carinata全基因组LAI得分为12.23,Bc和Cc亚基因组的LAI值分别为11.78和12.71,说明重复序列区域组装较为完整。
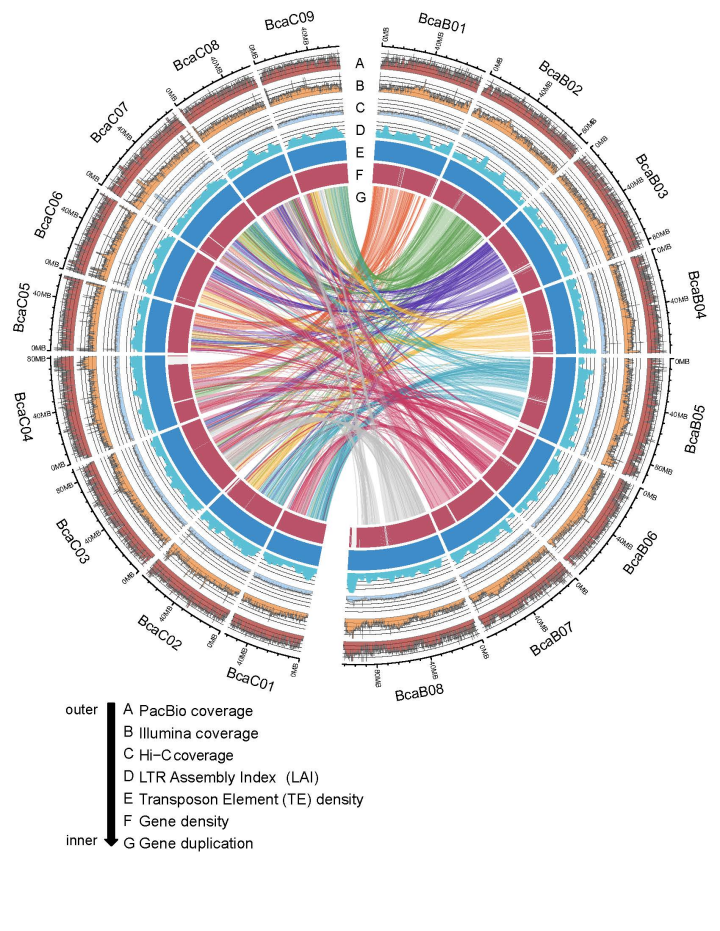
图2 Brassica carinata Gomenzer基因组组装的染色体特征
B. carinata基因组是禹氏三角异源四倍体物种中基因含量最高的,包含133,667个注释基因,预测产生143,117转录本(图3A),共有105,596个基因通过RNA-Seq比对得到支持,占全部注释基因模型的78.9%,其余基因通过Pfam结构域或BLAST同源性得到支持。B. carinata的每个亚基因组包含的基因数,比禹氏三角模型中各自共享基因组的基因数更多(图3D)。
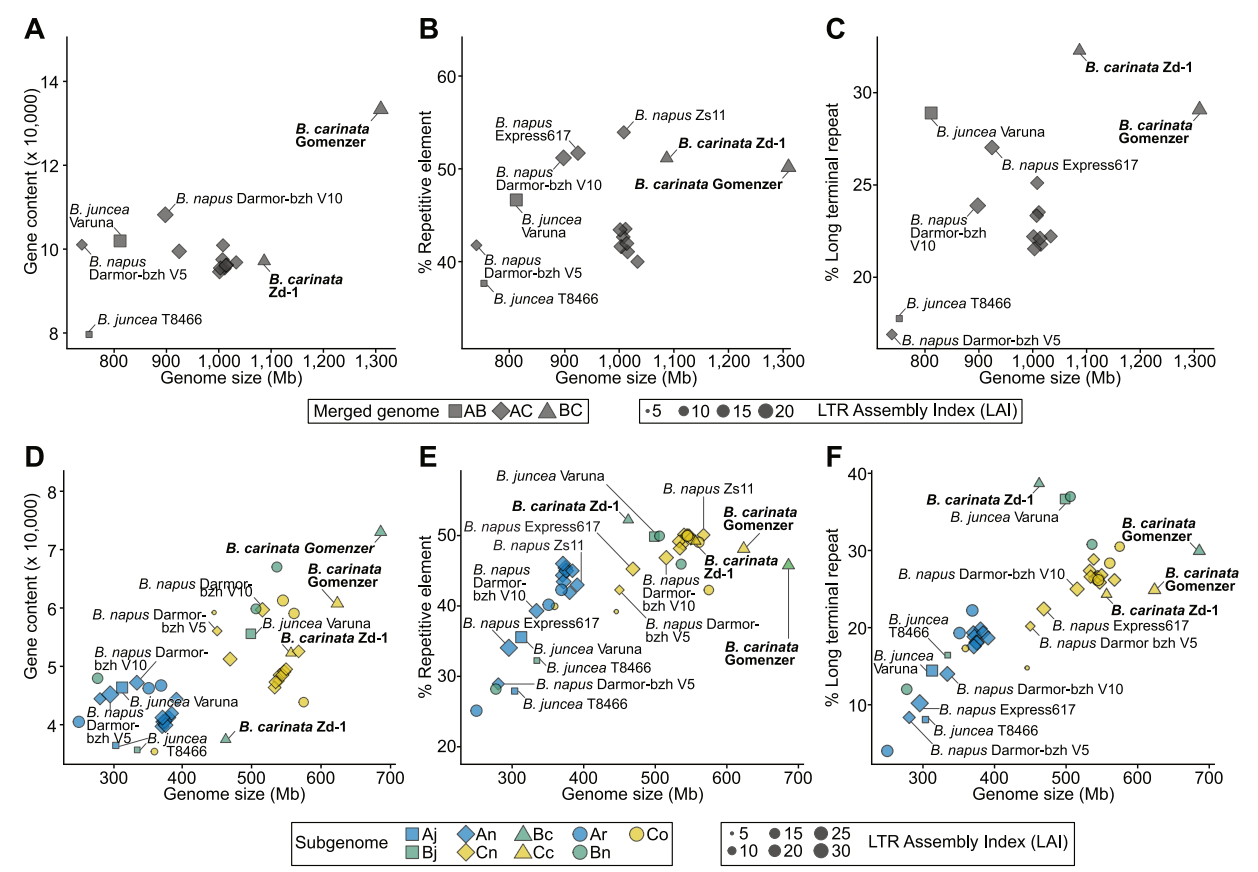
图3 芸苔属基因组大小和遗传特征比较
作者研究了B. carinata的基因分离(即同源亚基因组失去基因而回到二倍体状态的过程)。禹氏三角模型的基因组是高度重复的,二倍体自所有真核生物最近共同祖先以来,经历了总计36× multiplication(3×2×2×3),异源四倍体经历了总计72×的multiplication (3×2×2×3×2)(图4)。
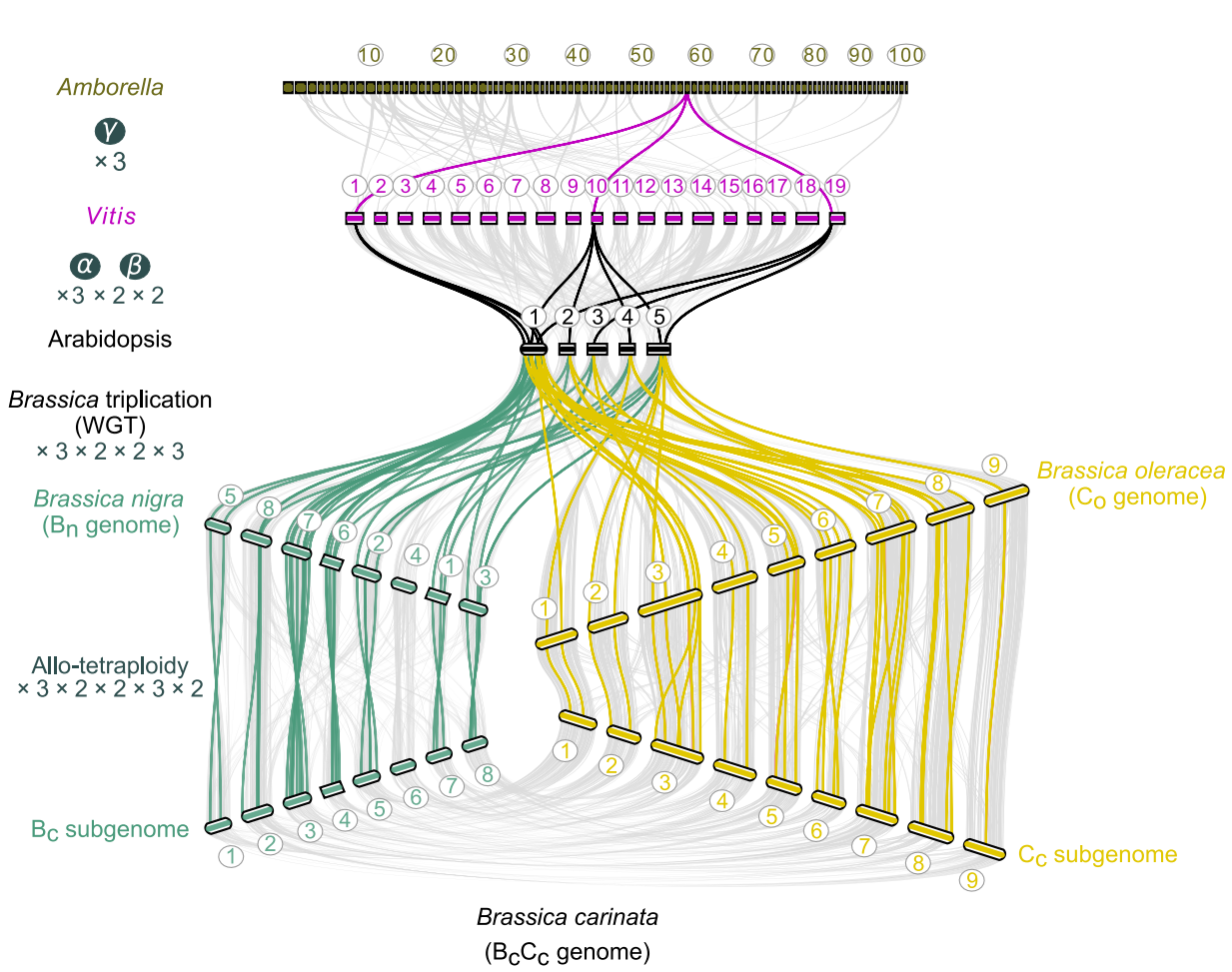
图4 支持芸苔属物种WGD事件的部分共线区域的展示
尽管B. carinata基因组经历了一定程度的二倍体化,但全基因组比较并没有显示大量的基因丢失。研究发现Bc亚基因组78.3%的基因来源于WGD,而Cc亚基因组中只有49.85%的基因来源于WGD(图4),与在植物中观察到的许多其他古多倍体化事件类似,B. carinata的重复基因明显保留。
2、 禹氏三角模型杂交事件时间预估
根据Ks分布图 (图5B),推断B. carinata物种形成的杂交事件(B. nigra(Bn)和B. oleracea (Co)之间杂交)发生在大约11000 - 29900年前,比此前研究估计的时间更近。
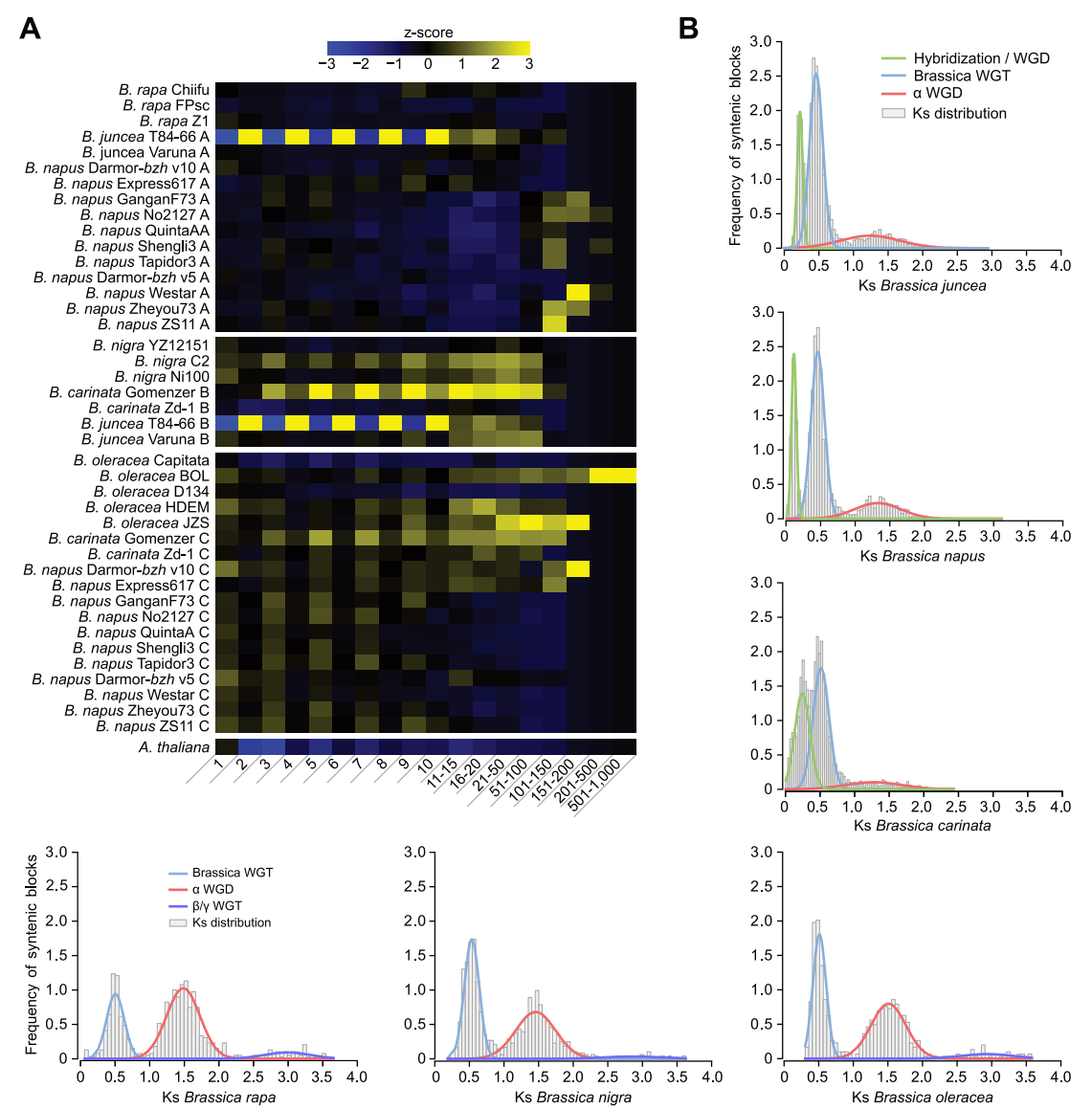
图5 基因家族与WGD事件
随后,作者构建了禹氏三角基因组的系统发育树,以解析Bc和Cc亚基因组的起源(图6A),分析发现Bc亚基因组从B. juncea的Bj亚基因组在~5.44 MYA分化而来,两者最后的共同祖先从B. nigra(Bn)基因组~6.61 MYA分化而来(图6A)。这些结果表明,B. carinata(BcCc)和B. juncea(AjBj)来自同一个B谱系。
另一方面,Cc亚基因组从B. oleracea(Co)甘蓝型谱系(BOL)中在2.88 MYA分化出来,B. napus(AnCn)的Cn亚基因组来自于甘蓝型B. oleracea(Co)谱系,其最近的分化估计约为2.63 MYA(图6A)。因此,B. carinata(BcCc)和B. napus(AnCn)可能起源于不同的谱系。
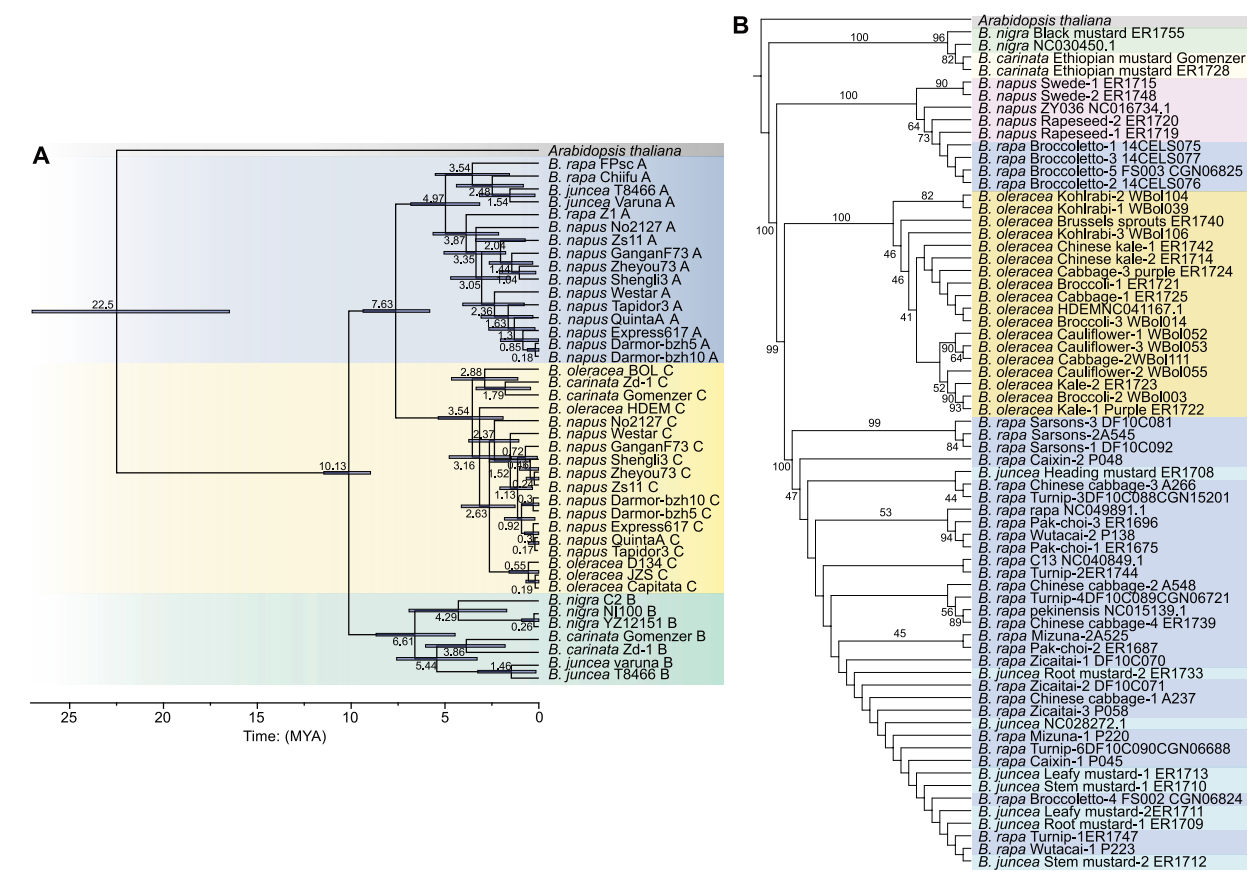
图6 禹氏三角基因组的进化关系与分歧时间估计
此外,通过B. nigra(Bn)和B. carinata(BcCc)组成的独特分支的鉴定,可以将B. nigra作为母系祖先(图6B)。类似地,B. napus(AnCn)和B. juncea(AjBj)与B. rapa(Ar)独立聚类,可以将B. rapa指定为这两个物种的母系祖先,这些观察结果与此前报道一致,认为B. rapa(ssp. Rapa)是B. napus的祖先。
3、TEs扩张与基因组的大小相关
TEs是植物基因组的重要组成部分,在植物基因组的结构和功能进化中起着至关重要的作用。B. carinata Gomenzer基因组在禹氏三角模型中包含的重复序列最多,且其两个亚基因组包含的重复序列都比它们各自的共享基因组(Bn, Bj, Co, Cn)包含的重复序列多。
在Bc和Cc亚基因组之间重复序列也存在不对称分布的情况。虽然两个亚基因组TE含量相似(Bc: 47.3%, Cc: 48.7%),但TE在染色体上的分布差异显著。Bc亚基因组中的TEs表现出典型的着丝粒周围富集和端粒富集,而Cc基因组中的TEs则更均匀的分散在不同染色体上(图7A, 7B)。

图7 Brassica carinata染色体的基因组图谱
此外,对B. carinata Gomenzer和B. carinata Zd-1组装结果比较显示,在禹氏三角模型的异源四倍体中,B. carinata基因组的平均LTR含量最高(图3C),且LTR含量不对称。Bc和Cc亚基因组中LTR序列分别占的29.9%和26.0%,进一步分析发现,在其他两个异源四倍体B. napus(AnCn)和B. juncea(AjBj)也中存在LTR的不对称分布。
4、基因家族扩张促进B. carinata驯化过程中的适应
该研究发现Bc亚基因组比Cc亚基因组包含更多的基因(图3D)。两个亚基因组的基因数量都高于禹氏三角模型基因组的平均水平,这表明基因家族的基因丢失较少,可能存在基因扩张。
与其他芸苔属植物相比,B. carinata的连续重复基因在多个通路上保留,可能是其提高环境适应性和新性状产生的根本原因。对扩张基因家族基因进行GO和KEGG分析发现扩张基因在光合作用、激素信号转导、植物-病原体相互作用、二萜类生物合成、抗坏血酸和醛缩酸代谢等通路的富集。表明B. carinata的这些基因和其他涉及重要农业性状的基因可以作为未来分子育种工作的潜在目标,以提高其他作物,特别是其他禹氏三角模型中物种的气候适应能力。
5、亚基因组偏倚研究
由于B. carinata基因组可能处于二倍体化的早期阶段,其82.6%的基因与至少一个各自的祖先基因组保持共线(Bn和Co,图8)。结合重复序列分析中得到的结果,表明Bc亚基因组比Cc亚基因组经历了更少的基因组重排,表现出高水平的分裂。
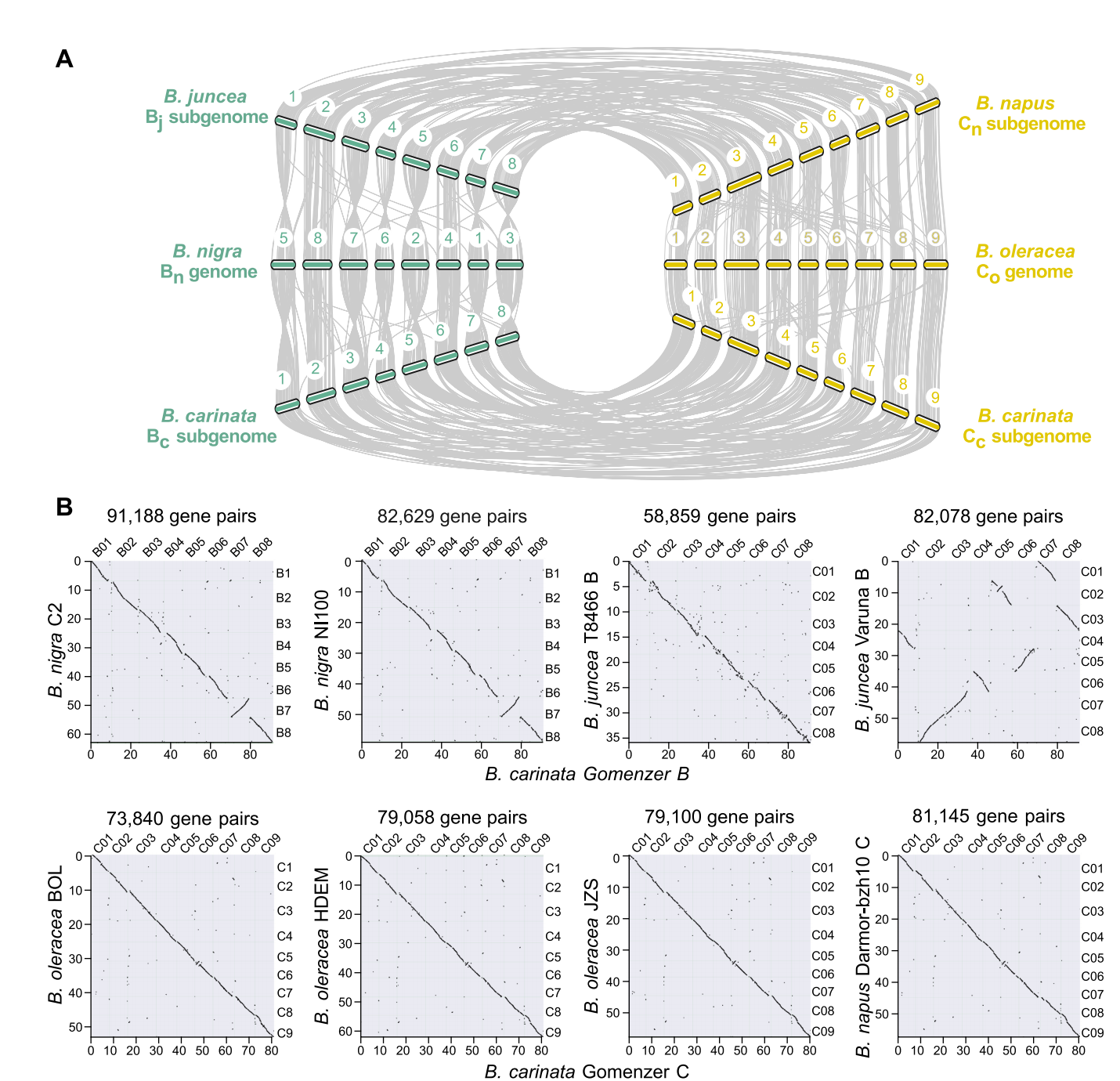
图8 芸苔属祖先(B. rapa,B. nigra和B. oleracea)以及杂交异源四倍体(B. carinata,B. juncea和B. napus)的基因组共线性分析
此外,一个亚基因组同源基因的表达高于另一个亚基因组,是亚基因组优势的标志之一。该研究使用RNA-Seq数据,比较共线同源基因对之间的基因表达(Bc与Cc)。大部分同源基因(60.4%)表现出相似的表达模式;22.6%的同源基因显示Bc亚基因组偏倚;18.3%的同源基因显示Cc亚基因组偏倚。此外,这种偏倚在所有类型的组织中都比较明显,在花组织(Bc: 22.1%, Cc: 16.7%)和热响应期间(Bc: 23.44%, Cc: 18.1%)最为明显。
该研究对比了B. carinata的序列进化速率,发现Cc同源异构体的平均ω =Ka/Ks更高,在RNA-Seq样本中,Cc亚基因组的ω=Ka/Ks值仍然较高。因此,研究结果表明,与Bc亚基因组相比,Cc亚基因组受到更弱的纯化选择压力,或更高的适应速率。
6、编码脂肪酸合成相关关键酶的基因研究
作者针对的重要农艺性状基因扩张和转录丰度进行研究,发现在B. carinata中,脂肪酸(FA)生物合成和延伸途径的基因发生了显著扩张。在高等植物中,脂肪酸的从头合成通路如图9A所示。
进一步研究发现Bc亚基因组优势可能会对生物功能和表型性状产生影响,与种子油生物合成和开花时间相关的关键基因,表现为Bc亚基因组偏倚表达(图9)。例如ACP由ACP1编码,在Bc亚基因组中存在8个拷贝,在Cc亚基因组中只有1个拷贝,Bc亚基因组多拷贝贡献了角果中98.4%的ACP1转录本,该研究结果为B. carinata分子育种提供重要基础。
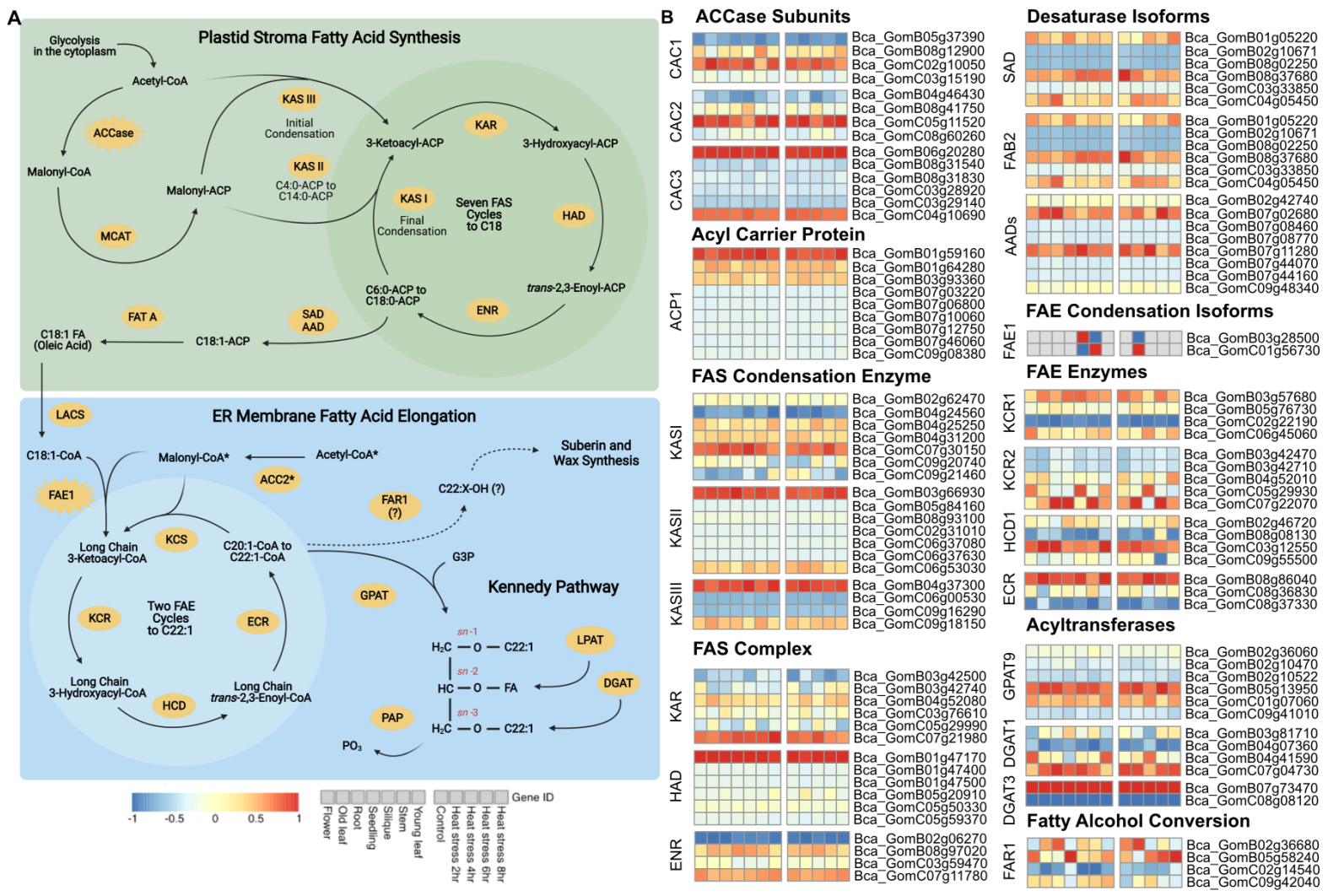
图9 芥酸三酰基甘油生物合成途径
总结
该研究组装的B. carinata基因组填补了禹氏三角模型中最后一块空缺,为生物燃料原料和多倍体进化研究提供新思路。以B. carinata基因组基础进行抗性相关基因研究,为其他十字花科蔬菜的优良性状分子育种提供了重要基础。
参考文献:
Yim WC, et al. The final piece of the Triangle of U: Evolution of the tetraploid Brassica carinata genome. Plant Cell. 2022.
AG代理基因使用Nanopore平台完成了全球首个大型复杂植物基因组菊花基因组的组装和后续分析工作。现已完成500余例真核生物的Nanopore及PacBio基因组测序及组装,服务项目包括:哺乳动物、鱼类、农作物、药用植物、昆虫、原虫、真菌等多类型复杂样本。由AG代理基因主导提出并推动千种本草基因组计划,并构建药用植物基因组数据库,已发表多篇药用植物基因组高水平文章,引领药用植物基因组研究前沿。


