 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
文献解读 | 园艺学顶刊Horticulture Research发表杜鹃gapless基因组及花色差异的多组学研究成果
杜鹃花属(Rhododendron)的花朵颜色高度多样化,为花色研究提供了独特研究模型。该研究用ONT测序数据对黄花杜鹃R. molle进行了gapless基因组组装,选用黄花/红花杜鹃进行比较基因组分析,结合代谢组、时序基因共表达等分析,揭示了不同花色素沉着的特定基因调控变化机制,为杜鹃花属的遗传学和分子育种提供重要基础。

文章题目:Gapless genome assembly of azalea and multi-omics investigation into divergence between two species with distinct flower color
发表期刊: Horticulture Research(IF= 7.291)
发表时间:2022.10.26
主要研究结果
1、R. molle的gapless基因组组装与注释
该研究使用51.97Gb(~100×)的ONT(Oxford Nanopore Technologies)测序数据,49.12 Gb的PCR-free Illumina数据,结合193.78 Gb的Hi-C数据进行R. molle染色体水平基因组组装。最终组装大小为653.46 Mb,获得了一个端粒到端粒的gapless基因组组装,染色体挂载率为99.48%,contig N50为44.85 Mb,BUSCO为93.4%,与已发表的多个杜鹃属植物基因组相比(表1),在连续性和完整性方面均有较大提升。
表1 9种杜鹃花植物的测序、组装和注释比较
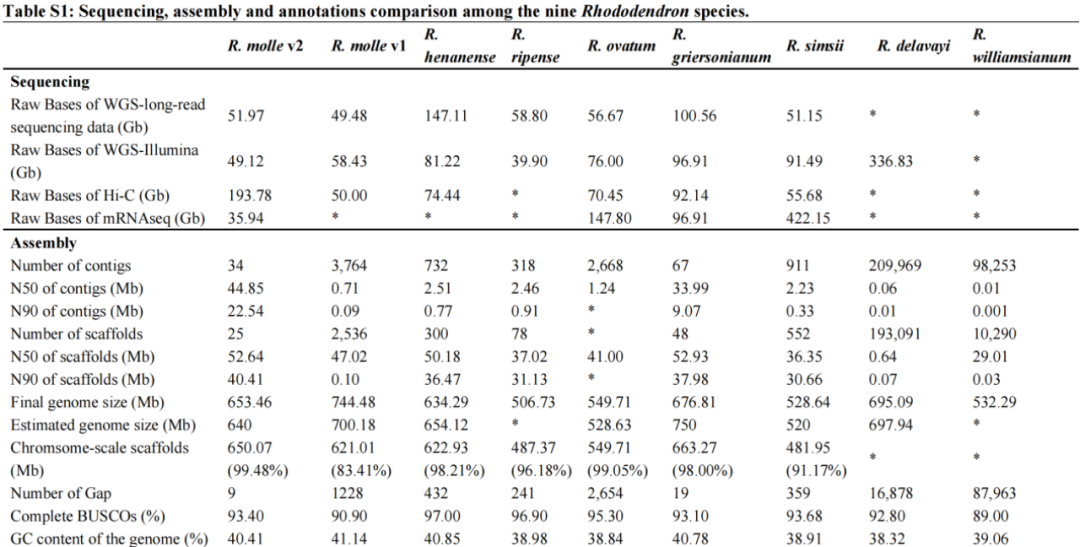
R. molle的13条假染色体各有一个着丝粒,着丝粒长度从3 Mb到20 Mb不等。并且Gypsy LTRs优先富集在着丝粒区域,从着丝粒向染色体末端延伸,GC含量下降,基因密度增加(图1a)。通过对端粒特征motif进行检测,在组装的染色体上共检测到17个潜在的端粒区域。其中,chr07和08完全为gap-free组装,且检测出两端的端粒,即达到T2T(telomere to telomere)水平。全基因组中仅有6条染色体存在9个gaps,其中4个gaps位于chr11非常复杂的着丝粒区域(图1a)。
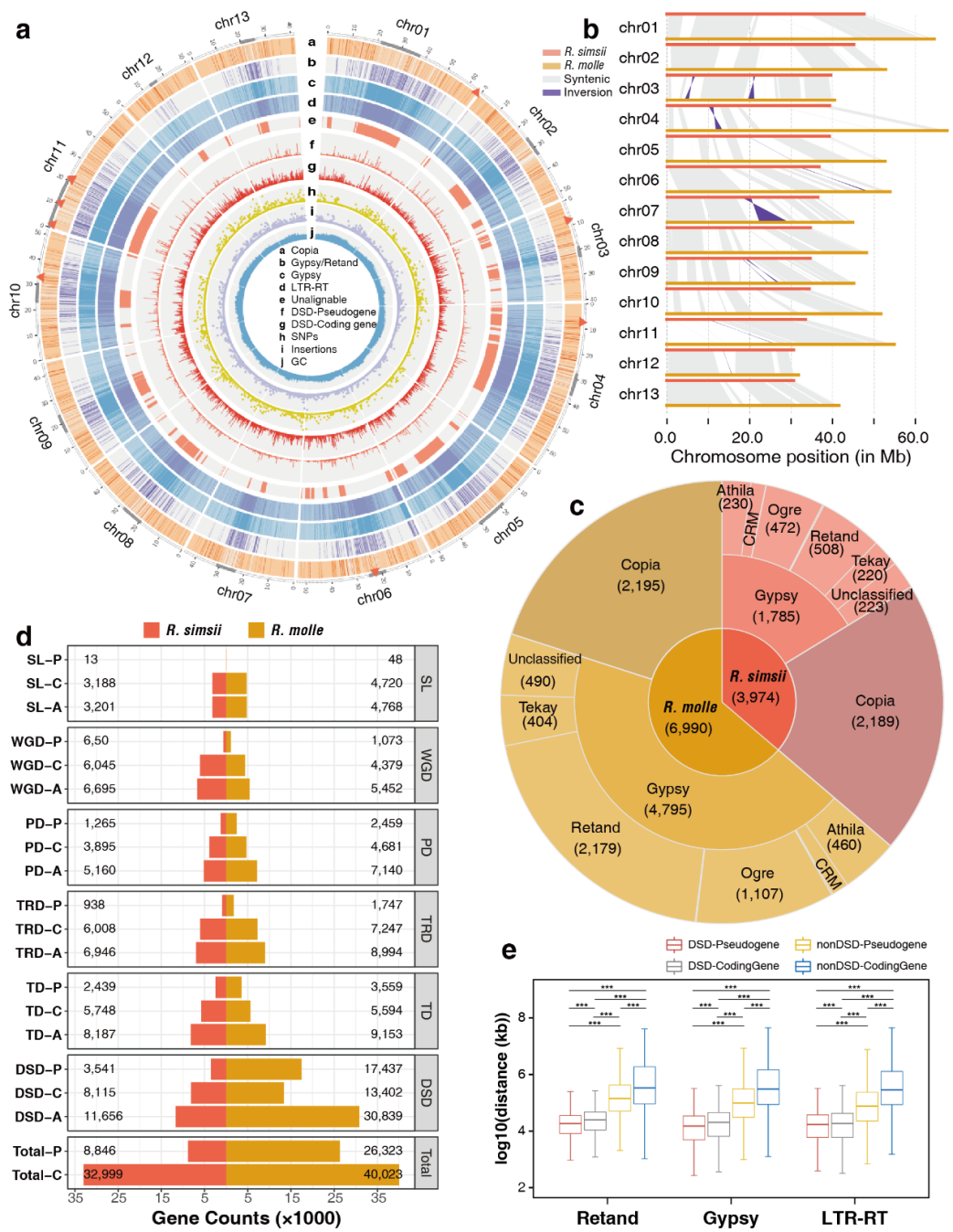
图1 R. simsii和R. molle的基因组比较
2、杜鹃的基因组差异和假基因化
共线性分析对R. simsii和R. molle基因组进行结构变异鉴定,除402,253,777 bp长的共线区(~占整个R. molle基因组的60%)外,有234,864,123 bp(~36%)是R. molle特有的,因此与参考基因组未对齐(图1b),这些无法对齐的区域是R. molle基因组中的LTR-RT富集区(图1a中a-e)。表明LTR-RTs可能导致了种间基因组的分化。有趣的是,作者发现大多数R. molle的特有序列(与R. simsii无法对齐的区域)都是潜在的着丝粒区域(图1a),表明这两个物种之间的着丝粒差异可能与物种分化相关。
LTR-RT和Gypsy的扩张与大量的基因复制和假基因化相关。与R. simsii相比,R. molle基因组中Gypsy的扩张主要是由于四个亚组的扩张:Retand、Ogre、Athila和Tekay(图1c)。该研究在R. molle中鉴定出26323个假基因和40023个编码基因,其数量远高于R. simsii(假基因8846个;编码基因32999个)(图1d)。在R. molle的编码基因和假基因中,散在重复(DSD)最多,有30839个(46.5%)。
3、代谢组和色素的生物合成
该研究重建了与R. molle和R. simsii花色形成有关的色素代谢途径(叶绿素、类胡萝卜素、花青素和黄酮醇等)(图2、3a)。
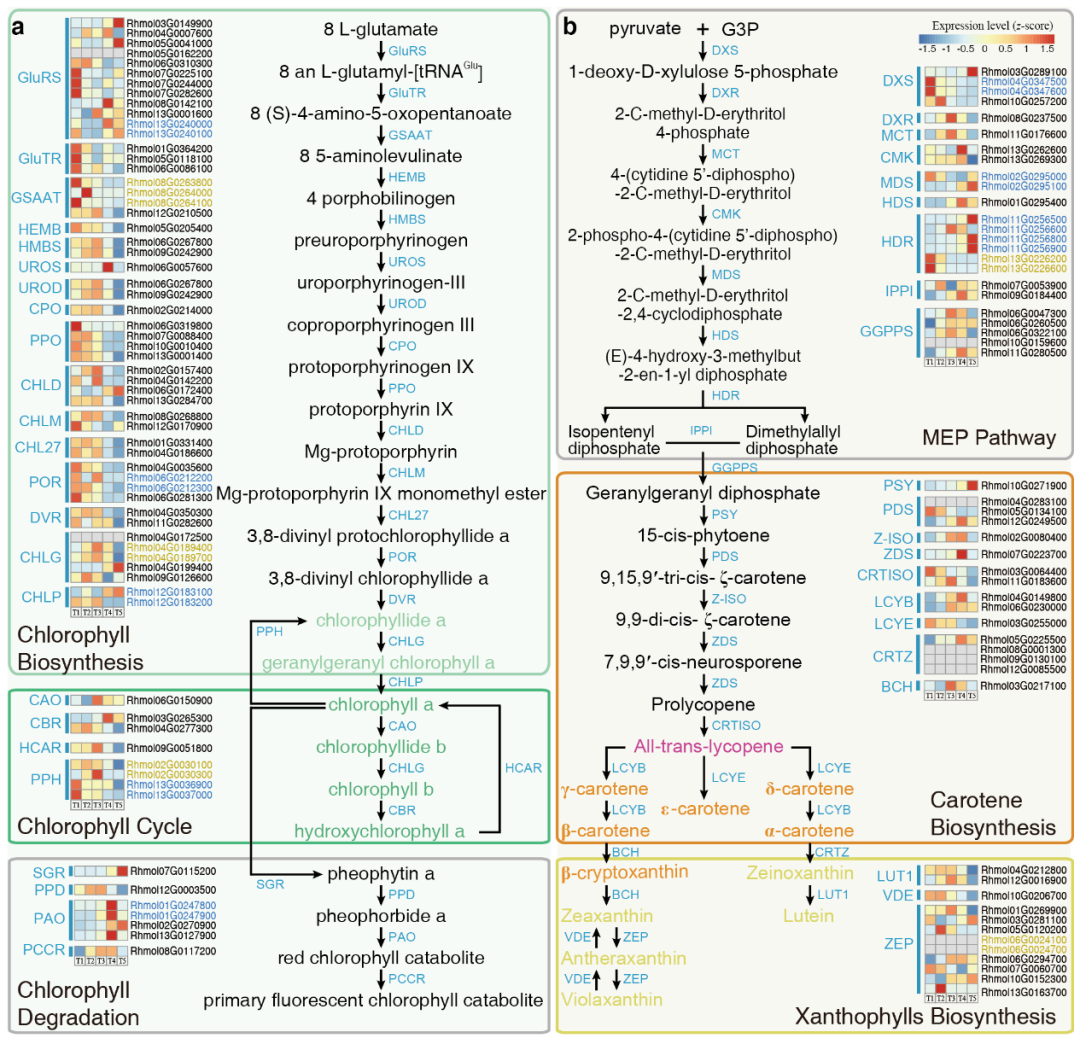
图2 R. molle类胡萝卜素和叶绿素的代谢途径和时序基因调控
作者比较了R. molle和R. simsii基因家族,发现15个花青素生物合成途径中有12个基因家族发生收缩,其收缩程度(80%)在R. molle的所有生物合成途径中最高(图3b)。在R. molle花青素合成的基因家族中也发现较多假基因,表明假基因化可能是导致该物种中观察到基因家族收缩的原因(图3c)。
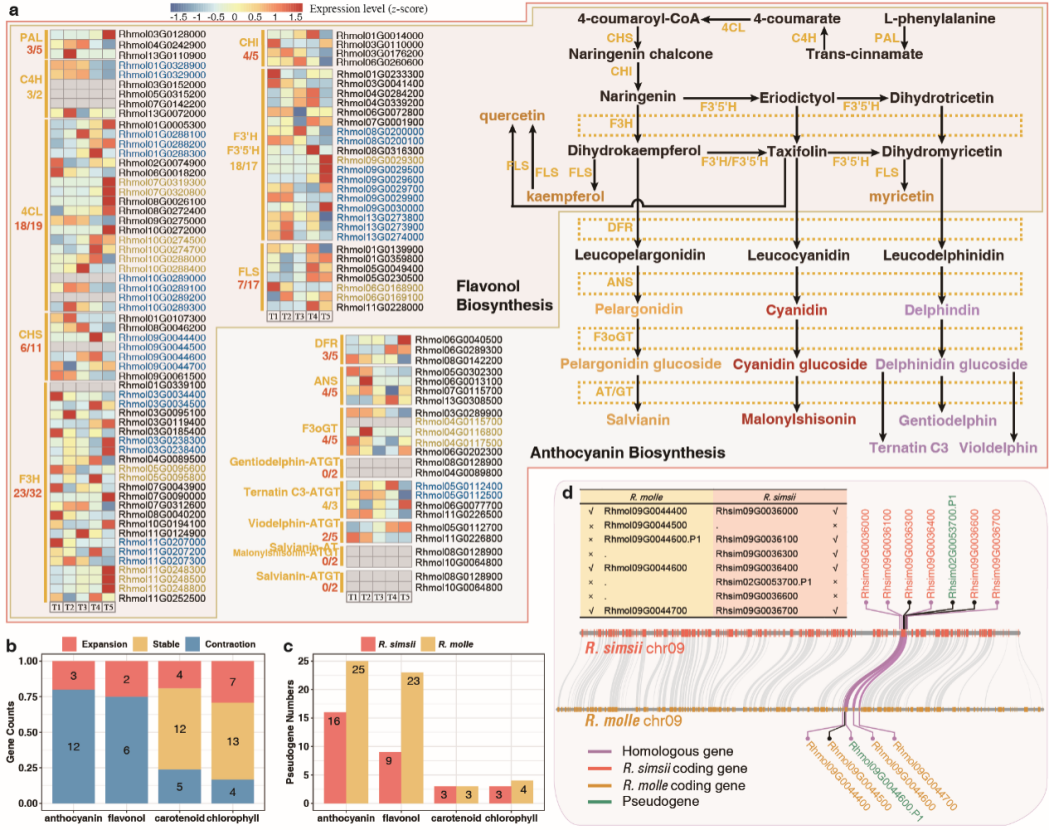
图3 花青素生物合成基因的缺失导致了R. molle中缺少红色品种
在叶绿素和类胡萝卜素的代谢途径中,收缩的基因家族和假基因相对罕见(图3b,c)。由于黄酮醇与花青素有7个生物合成步骤(图3a),在黄酮醇途径中也发现了相当数量的假基因和收缩的基因家族(图3b,c)。图3d为类黄酮生物合成通路基因获得和缺失的示例。
4、不同杜鹃之间时序基因共表达的比较分析
该研究分别构建并比较了两种杜鹃花的时序基因共表达网络(TO-GCNs),研究了两种杜鹃花之间与花色变化相关的基因调控(图4)。
根据花色素沉着的表达模式,时序子网络可分为三个主要阶段:初期(T1-T2,L1-L3,花瓣仍为绿色)、中期(T3,L4,绿色到黄色过渡)和末期(T4-T5,L5-L7,黄色)(图4d)。
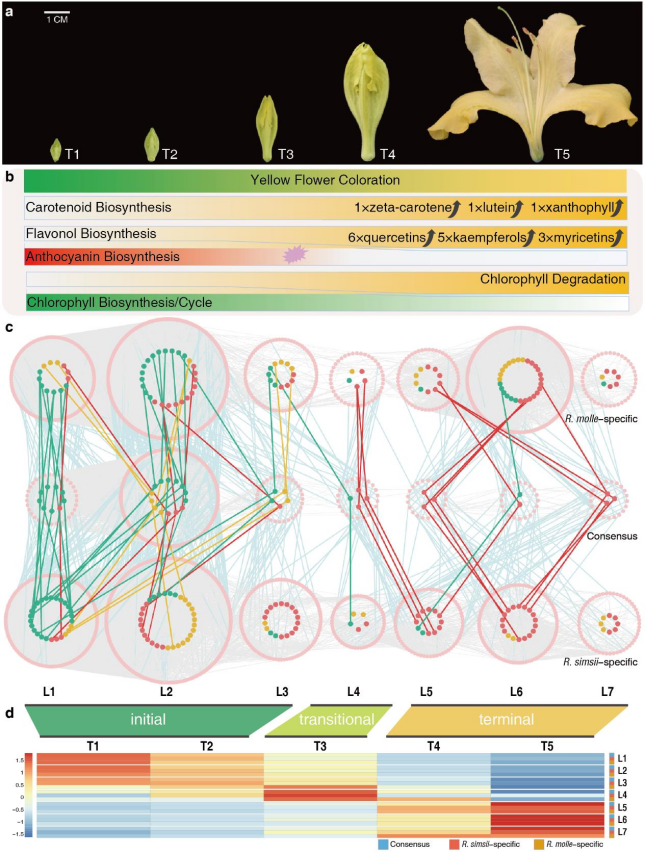
图4与黄花颜色相关的色素沉着变化和时序基因共表达网络
在检测物种特异基因数量时,两种杜鹃花的末期差异最为显著。两种杜鹃花在开花初期均表现出保守的共表达,包括239个转录因子和31个与色素生物合成相关的酶促基因(图5a)。在初期阶段,叶绿素生物合成基因是酶促基因中最丰富的类型,其次是类胡萝卜素生物合成基因、花青素生物合成基因和黄酮醇生物合成基因。在末期,两种杜鹃花之间的TO-GCNs的有限一致性(图5a)有助于物种特异性色素调控的鉴定。
进一步分析发现,类胡萝卜素、黄酮醇类胡萝卜素、黄酮醇生物合成基因在黄花R. molle中亚网络中被识别,而花青素生物合成基因位于红花R. simsii亚网络中被识别,进一步证实了代谢组研究结果。
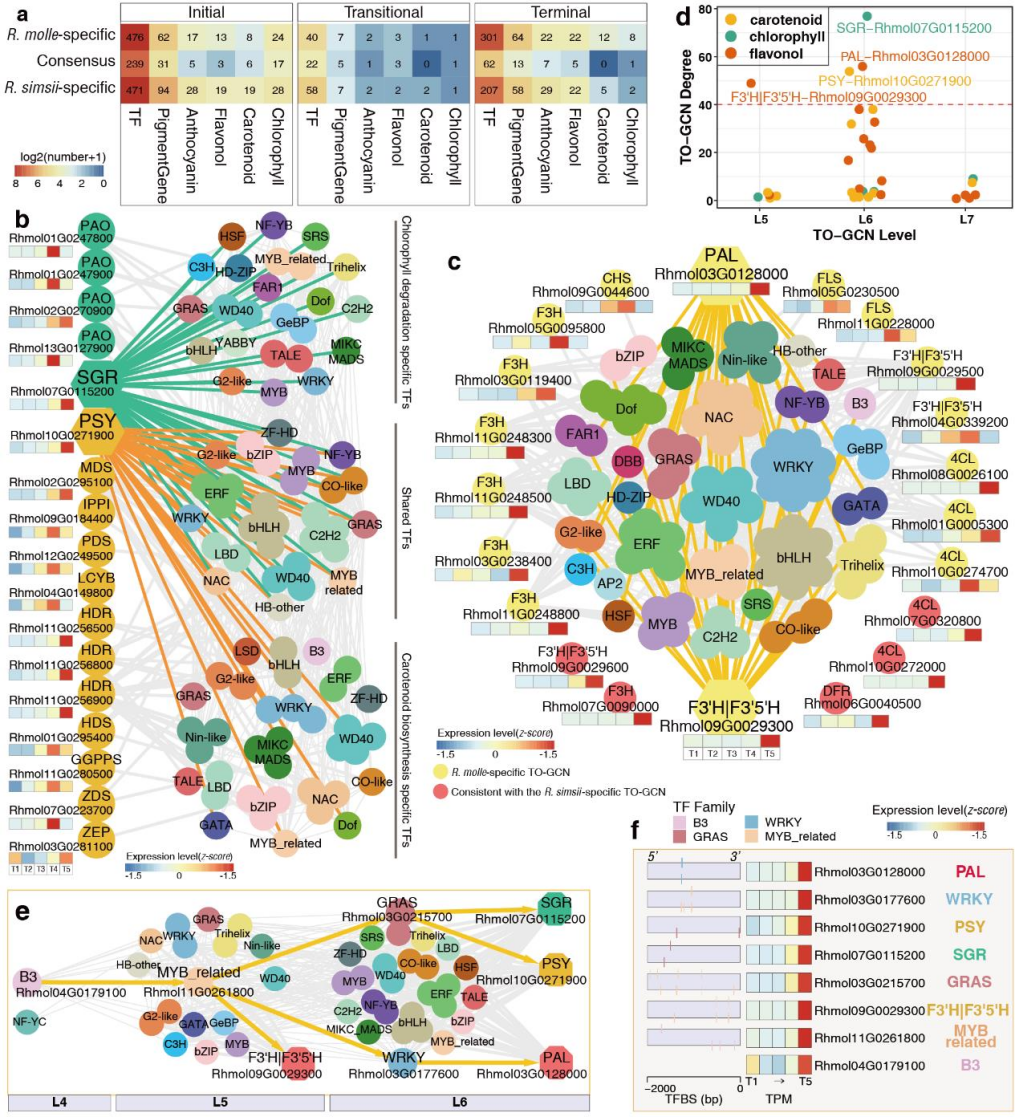
图5 R. molle特异性TO-GCN末期色素代谢途径的子网络
在R.simsii特异性亚网络中,作者鉴定出19个主要产生花青素的酶基因,这些基因均在末期高水平表达(图6a),预测这些酶基因可能受到74个潜在调控因子的调控,主要是WRKY、ERF、WD40、C2H2和NAC家族成员。有意思的是,在R.simsii特异亚网络95个基因的103个假定的同源物与R. molle中LTR-RT插入显著相关(图6b,c)。
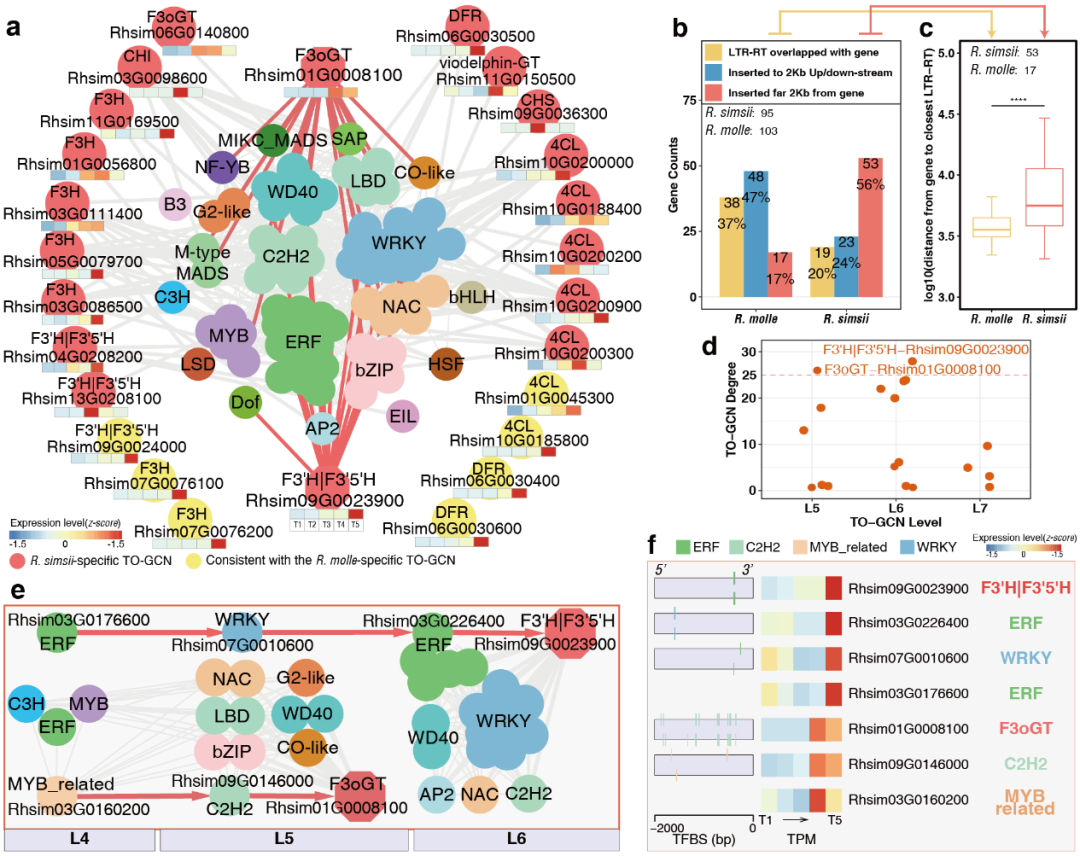
图6 R.simsii特异性TO-GCN末期色素代谢途径的子网络
作者进一步检测了hub gene F3oGT 和F3'H|F3'5'H(图6d)及其35个潜在调控因子(图6e)。发现F3oGT可能由C2H2和MYB_related共同调控;F3H|F3‘5’H可能由ERF,WRKY和ERF共同调控(图6f)。
总结
该研究利用ONT数据,结合Illumina和HiC测序技术,组装了gapless的黄花杜鹃(R. molle)高质量基因组,多条染色体达到T2T水平。通过黄花/红花杜鹃的比较基因组分析,发现Gypsy扩张与基因组大小的增加及散在重复、假基因数量的增加相关。结合代谢组、时序基因共表达网络(TO-GCNs)和比较分析揭示了不同花色素沉着的特定基因调控变化机制。
本研究采用多组学策略,解开两种重要杜鹃花之间的花色差异机制,为进一步的功能遗传学和分子育种提供参考。
参考文献:
Shuai Nie, et al. Gapless genome assembly of azalea and multi-omics investigation into divergence between two species with distinct flower color. Horticulture Research, 2022.


