 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
Poly(A)尾研究文献解读|TOP mRNA poly(A)尾长和核糖体装载的mTOR和LARP1依赖性调节
编码蛋白质合成机制的5‘末端寡嘧啶(TOP)mRNA的翻译受到氨基酸传感mTOR途径的严格调控。然而,其调控机制仍然未知。
本研究通过直接RNA测序(dRNA-seq)等一系列实验证明LARP1有助于在氨基酸饥饿(AAS)下具有长poly(A)尾部的TOP mRNA的选择性积累,从而加速核糖体加载到TOP mRNAs上,以便在AAS后恢复翻译。

英文标题: mTOR- and LARP1-dependent regulation of TOP mRNA poly(A) tail and ribosome loading
发表时间:2022.10.25
发表期刊:Cell Reports
IF:9.995
研究背景
雷帕霉素靶蛋白(mTOR)是连接细胞代谢、mRNA翻译和最终细胞生长的营养传感通路的中心靶点。氨基酸可以诱导mTOR的激活,在转导mTOR活性改变的蛋白中,RNA binding蛋白LARP1在mTOR信号通路下游受严格调控的核糖体生物发生中起着至关重要的作用。这些LARP1功能在mTOR信号通路下游受到调控:当mTOR信号通路被AAS或mTORC抑制剂失活时,更多的TOP mRNA被LARP1结合,变得更加稳定,但翻译被抑制。
Poly(A)尾部代谢与基因表达密切相关。poly(A)尾部去除,称为去烯基化,是mRNA衰变的第一步和限速步骤,去烯基通常是被序列特异性RNA结合蛋白靶向以破坏mRNA的稳定性。相反,在一些种类丰富的mRNA中,去烯基化和mRNA降解可以解耦。然而,poly(A)长与翻译的解耦是否适用于所有种类的mRNA,是否有例外,LARP1介导的poly(A)尾长调控的意义可能是什么,目前尚不清楚。
材料方法
材料:HEK293T细胞
主要研究策略:direct RNA sequencing、Northern blotting
研究结果
1、 在富含氨基酸的条件下,TOP mRNA翻译与其poly(A)尾部长度呈正相关
为了研究poly(A)尾巴长度与翻译之间的相关性,研究者对细胞质提取物进行了多聚体分离,收集了4个馏分池,分别对应于游离(游离)、单体(单体)、轻多体(轻聚,2 - 4个结合核糖体)和重多体(重聚,5个以上结合核糖体)馏分(图1A)。使用帽结合蛋白GST-eIF4EK119A的高亲和力突变体从LiCl沉淀的总RNA中纯化5’帽mRNA,并进行基于dRNA-seq的poly(A)尾长分析,发现长poly(A)尾的密度在多体馏分中偏高(图1B)。
作者进一步找出poly(A)尾巴长度和核糖体密度之间呈现正相关的基因,GO分析表明,RP基因在正线性组中显著富集(图1D)。几乎所有的RP基因在其最末端都有5‘TOP序列。
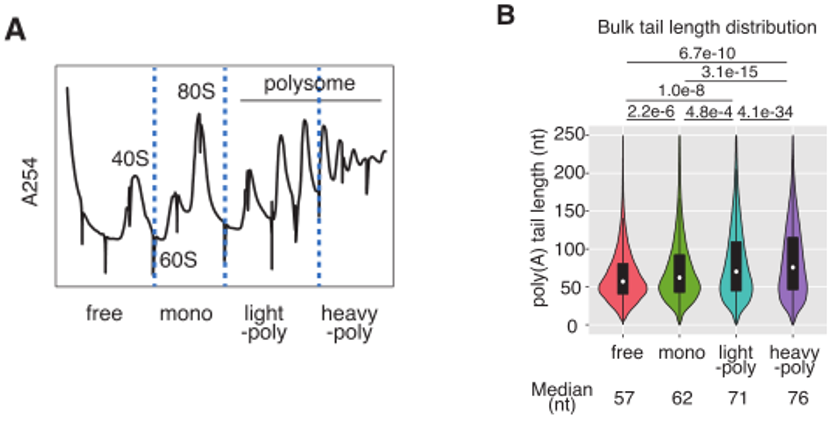
图1在TOP基因中,翻译和poly(A)尾巴长度的耦合作用很明显
Northern印迹分析显示,具有长poly(A)尾的报告mRNA(3h)主要分布在重多聚体部分,并且随着去烯基化的进行(6和9h),报告mRNA的分布向较轻多体部分转移(图2C-D)。
图2 dRNA-seq结果的Northern印迹验证
2、 TOP mRNA的Poly(A)尾长度随着氨基酸可用性的变化而动态波动,这种波动依赖于LARP1
在mTOR活性/富含氨基酸的条件下,TOP mRNA翻译和poly(A)尾部长度之间的相关性可能表明TOP基因在转录后受poly(A)尾部长度调控。因此,作者首先通过northern印迹检测了长期AAS对RP mRNA poly(A)尾部长度的影响(图3A)。迁移速率较慢的RP mRNA在AAS期间开始不断积累,而在生长条件下迁移速度更快的RP mRNA逐渐减少(图3A)。通过寡聚(dT)/RNase H消化去除poly(A)尾部,在整个样品的相同位置呈现单个条带,表明迁移较慢是由于尾部延长(图3A)。这些mRNA的poly(A)尾部长度响应于氨基酸的可用性而动态调节,因为当参考饥饿细胞时,拉长的尾部再次缩短(图3B)。
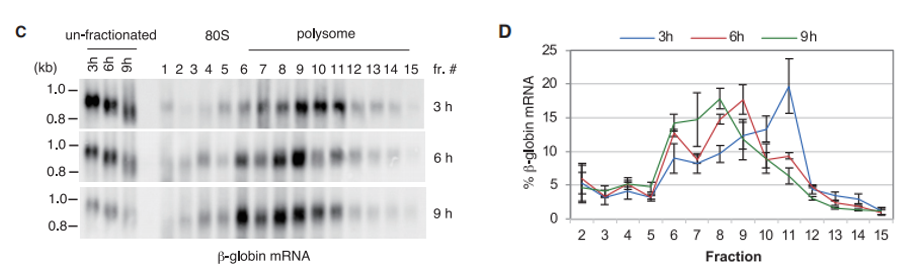
图3 AAS条件下的Northern印迹分析
为了获得AAS或mTOR抑制条件下与LARP1 KD结合的poly(A)尾长度变化的全貌,研究者进行了DRS分析,发现TOP mRNA的poly(A)尾部在Torin1处理下以LARP1依赖性方式延长(图4)。表明TOP mRNA poly(A)尾部在AAS/mTOR抑制条件下选择性延长,LARP1对该过程至关重要。
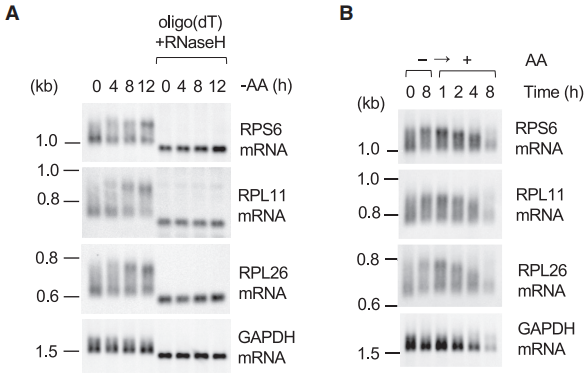
图4 mTOR失活下TOP mRNA的选择性和LARP1依赖性尾部延长
3、 LARP1介导靶mRNA转录后聚腺苷酸化
为了探索LARP1如何诱导靶TOP mRNA的poly(A)尾部延长,我们采用了MS2系留系统。在HEK293T细胞中,与MS2外壳蛋白N端融合的LARP1与携带噬菌体MS2干环的报告mRNA共表达,并通过northern印迹法评估mRNA poly(A)尾部长度。LARP1的系带导致报告mRNA的迁移较慢(图5A-B)。当去烯基化被抑制时,poly(A)尾部变得更长,这表明LARP1系链引起的poly(A)尾部长度的变化不仅仅是由于去烯基的抑制。
进而监测转录关闭后的poly(A)尾部代谢,在去烯基化缺陷条件下(HA-Caf1 mt/Pan2 mt)的LARP1系链导致在转录关闭后出现更长的poly(A) 尾mRNA,表明 poly(A)尾伸长发生在转录后,额外的转录追逐实验(图5H-I)结果说明LARP1诱导靶mRNA的转录后聚腺苷酸化。
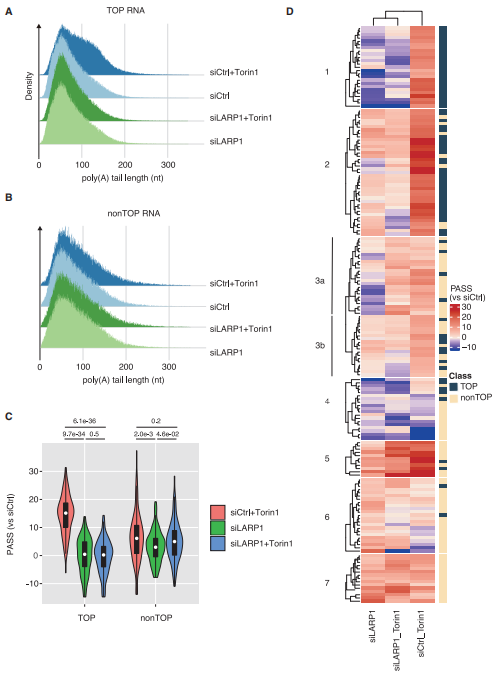
图5 LARP1转录后延长mRNA poly(A)尾部
4、 LARP1通过其N末端区域与非标准poly(A)聚合酶结合
最后,我们研究了LARP1是否与poly(A)聚合酶(PAP)发生物理相互作用。53FLAGLARP1过表达下的共免疫沉淀(coIP)实验揭示了PAPD4、PAPD5和PAPD7与LARP1的RNase抗性相互作用(图6A),PAPD4、PAPD5和PAPD7优先与LARP1的N端半部(1–495 aa)结合(图6C),这在与mRNA连接时触发poly(A)尾部伸长。
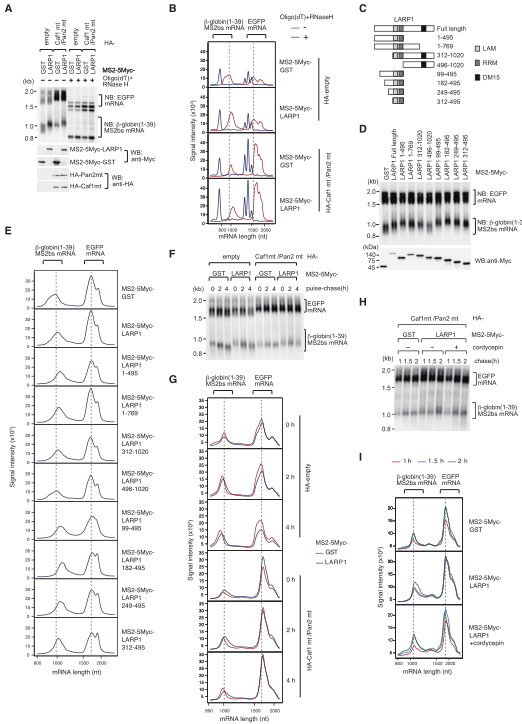
图6 LARP1与非典型poly(A)聚合酶PAPD4、PAPD5和PAPD7的RNase抗性相互作用
研究结论
本研究证明了在mTOR活性/富含氨基酸的条件下,TOP mRNA翻译与其poly(A)尾部长度呈正相关,这表明TOP mRNAs在转录后受poly(A)尾部长度调控。TOP mRNA的尾部长度随着氨基酸的可用性而动态波动。Poly(A)尾部在mTOR活性/富含氨基酸的条件下缩短,而长尾TOP mRNA在mTOR-非活性/氨基酸缺乏(AAS)条件下积累。一种RNA结合蛋白LARP1对该过程是必不可少的。LARP1与非标准poly(A)聚合酶相互作用,并诱导靶向的转录后聚腺苷酸化。


