 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
Nature顶刊详解|Direct RNA测序助力解析基因组守护者SAFB蛋白精准调控自主转座子的内在机制
英文标题:Autonomous transposons tune their sequences to ensure somatic suppression
发表时间:2024.1
发表期刊:Nature
IF:64.8
通讯作者:Tuğçe Aktaş(Max Planck Institute for Molecular Genetics)
研究介绍
转座子(Transposable elements,TE)是人类基因组中的重要组成部分,大约占据一半的内含子区域。在前体mRNA合成过程中,内含子中的转座子会与宿主基因一起进行转录,然而由于会与内含子一起被剪切和降解,它们大多数转座子序列最终无法成为成熟mRNA的一部分。矛盾的是,转座子同样也是RNA加工信号的重要来源,通过这些信号可能促使转座子形成新的内含子并产生不同的嵌合转录本。这种现象意味着细胞体内存在一种弹性剪切调控机制,可以抑制转座子外显子化,而不影响宿主前体mRNA加工。
基于前述,本文作者深入解析SAFB蛋白在mRNA加工过程中对转座子的调控作用,表明其可通过抑制转座子L1元件的逆转座来保护基因组的完整性,同时防止先前整合的转座子外显子化来维持剪切的完整性。其中,在本研究中,作者巧妙利用Nanopore Direct RNA测序可鉴定分析可变多聚腺苷酸化位点这一优势,对转座子的可变多聚腺苷酸化进行分析,证实潜在PAS位点的激活会影响SAFB上下游peak基因的差异表达以及PAS位点使用情况变化。
研究结果
1. SAFB蛋白结合转座子L1并抑制其逆转座
为了确定基因组中调控转座子剪切的作用元件,使用FLASH技术在HEK293细胞中鉴定到33个RNA结合蛋白(RNA-binding protein,RBP)的结合位点,随后进行UMAP降维聚类,发现有两个相邻聚类cluster2和cluster0富含Alu逆转座子结合位点,并且两个聚类各自的反义Alu元件占比与peak蛋白来源存在差异。进一步分析每个聚类中的转座子富集情况,结果显示cluster1中正义L1转座子占比最多,其主要结合三种具有ER型重复序列的SR同源蛋白:SAFB1、SAFB2和SLTM,它们构成了哺乳动物中的SAFB蛋白家族(图1)。依据结合模式来分析三种蛋白的结构和序列,发现SAFB家族蛋白的靶向作用区域存在高度重叠(图2a),与前述UMAP降维聚类结果一致。同时还在结果中观察到罕见的正义Tigger转座子的富集(图2b),其反义Tigger转座子与反义L1转座子在SAFB蛋白peak中被特异性去除,表明两类转座子在正义链上的序列存在共享,而在反义链上则没有(图2c)。
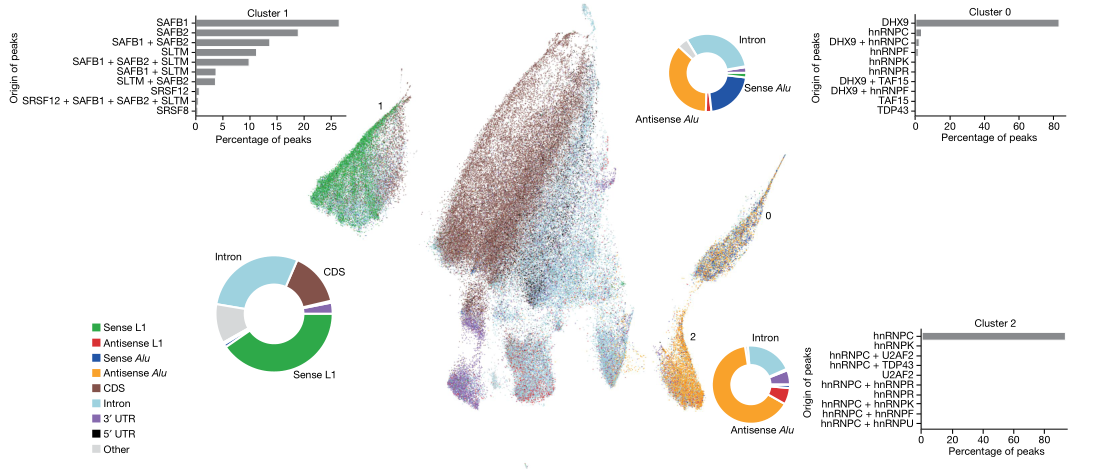
图1:HEK293细胞中RNA结合蛋白位点的UMAP降维聚类分析
作为基因组中的自主转座子,L1与结合蛋白的互作可能不仅仅影响宿主基因的RNA加工过程。三种SAFB蛋白均表现出对转座子中未片段化的、较长L1元件具有结合偏好性,并且结合区域主要集中在L1重复序列ORF1p和2p的编码区(图2d)。荧光素酶检测显示SAFB1的敲低会提高逆转座效率,而同时敲低三种SAFB蛋白时,L1逆转座效率提高更为显著,说明三种SAFB蛋白的功能可能存在冗余(图2e)。FISH原位杂交结果显示转座子L1的RNA仅位于细胞核内,而缺失SAFB1时,细胞质中L1的RNA含量则会显著增加,表明SAFB蛋白可结合转座子L1,维持其定位在核内,并抑制逆转座发生(图2f)。
哺乳动物中转座子L1 RNA的编码区富含腺嘌呤(A),并且可以通过优化密码子序列来调节腺嘌呤含量,比如在高度活跃的L1突变RNA ORFeus中,腺嘌呤含量明显降低。由此推测如果SAFB蛋白对转座子L1的抑制作用依赖于含有更多腺嘌呤的ORF,那么ORFeus经过优化后应当不受SAFB蛋白的调控影响。免疫印迹结果显示,L1转染细胞中单独敲除SAFB或L1转录调节因子HUSH,会导致ORF1p表达水平相同程度的增加,并且同时敲除时,表达水平增加会更加明显(图2g),而敲除SAFB并不影响ORFeus表达,但敲除HUSH则会使ORFeus表达上调,这一点通过RNA blot也得到验证(图2h)。以上结果表明,哺乳动物中转座子可通过维持编码序列中的腺嘌呤偏好性,以受到SAFB蛋白的抑制调控。
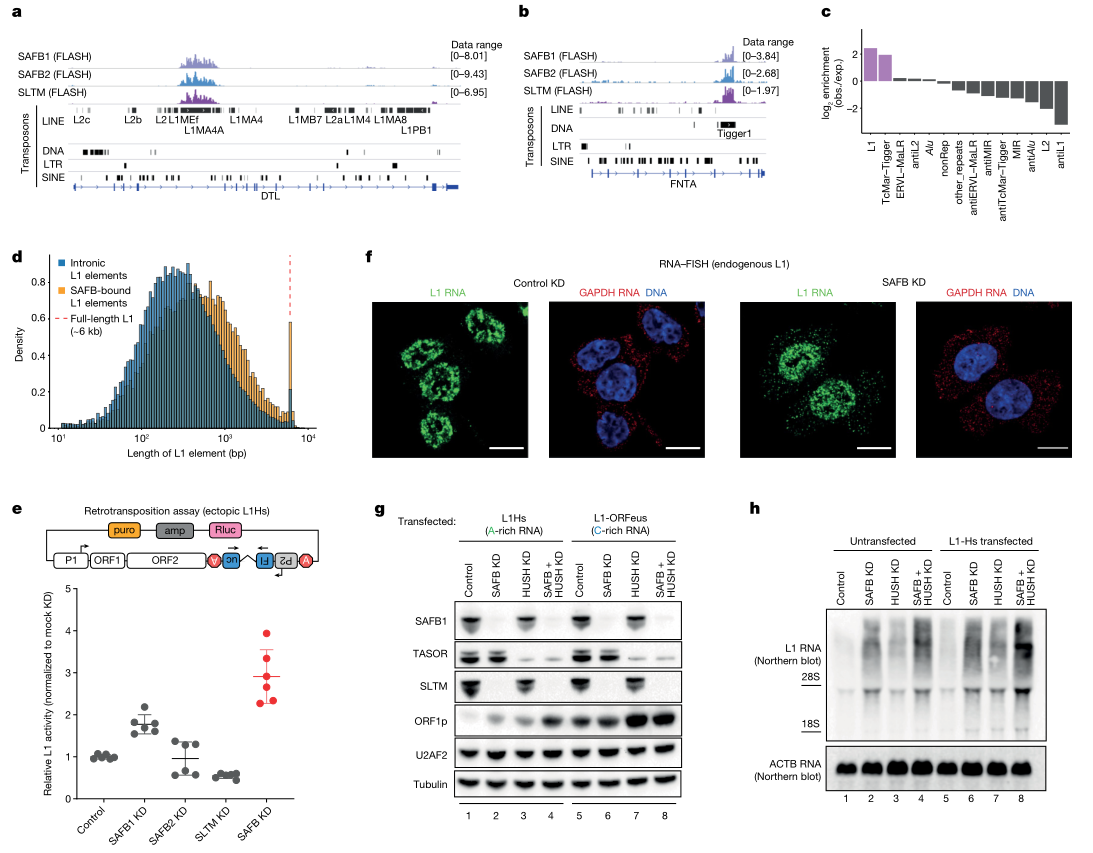
图2:SAFB蛋白结合转座子L1并抑制其逆转座
2. SAFB蛋白抑制转座子外显子化
转录组测序表明敲减SAFB家族蛋白会使得L1类和Tigger类转座子RNA表达水平呈现不同程度的上调(图3a),结合前述结果,说明SAFB蛋白可能参与转座子转录后调控。自主转座子L1或Tigger能够编码蛋白,理应存在多聚腺苷酸化(PAS)位点以形成具有翻译能力的 mRNA。正常情况下宿主RNA加工过程中可以跳过这些PAS位点,因此推测SAFB蛋白缺失可能激活潜在PAS位点,形成嵌合转录本,导致转座子RNA表达水平变化(图3b)。转座子在核内与胞质中的丰度增加表明嵌合转录本的形成可能与SAFB蛋白缺失引起的de novo剪切事件有关,这种de novo事件的主要特征是:与SAFB结合的转座子表达明显上调,因为PAS上游的至少一部分转座元件将会成为新的末端外显子的一部分;同时SAFB peak下游的外显子表达明显下调,因为在RNA聚合酶II到达这些外显子之前会终止转录(图3d)。对有无SAFB peak的基因及外显子在SAFB peak上下游的基因表达进行分析,发现宿主基因中SAFB peak的存在确实影响peak下游外显子的表达(图3c~d)。通过Nanopore Direct RNA测序对可变多聚腺苷酸分析,证实潜在PAS位点的激活是上下游peak基因差异表达的原因,并确定247个基因在SAFB缺失后PAS位点使用情况发生变化,同时约有5%的长转录本受到SAFB缺失影响而发生断裂。
SAFB蛋白缺失导致转座子潜在PAS位点激活,使原来位于内含子的PAS位点进入外显子区域,从而导致宿主基因的表达提前终止。L1和Tigger转座子均是无内含子的单外显子基因,在复制过程中并不需要经历剪切过程。然而,在缺失SAFB时,这两种转座子中都富含了可以作为剪切增强子的序列。FLASH结果表明SAFB蛋白可以识别一种外显子剪切增强子motif,典型特征是序列富含腺嘌呤,该motif可促进无内含子的RNA分子滞留于核内。转座子L1和Tigger恰好同样呈现出这一特性。
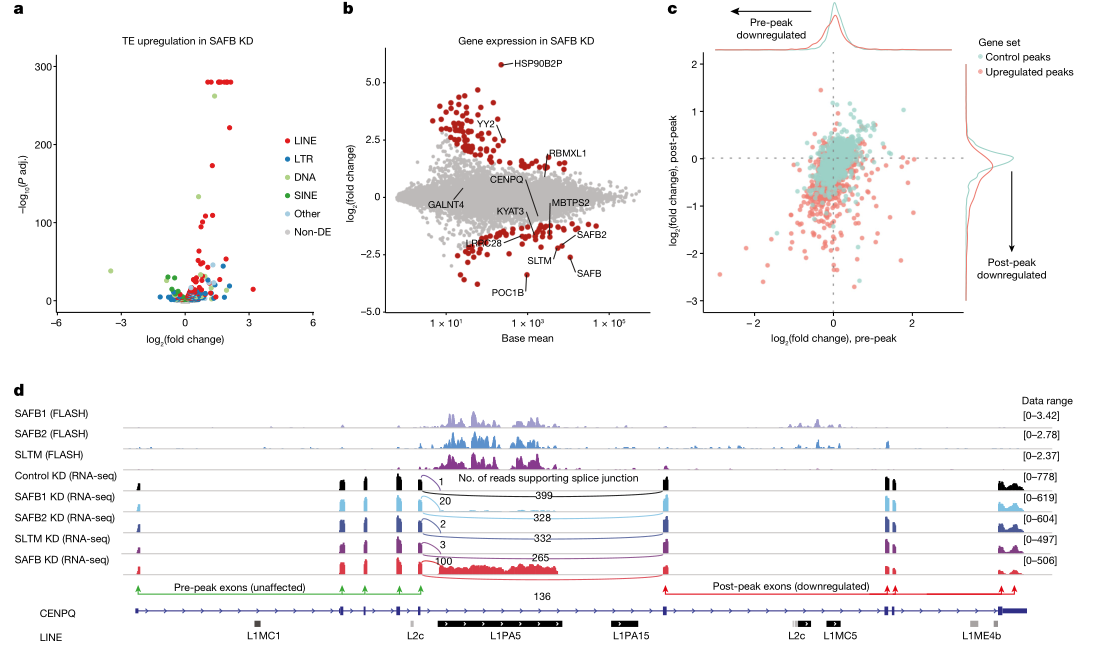
图3:SAFB蛋白抑制转座子外显子化
3. m6A修饰介导SAFB蛋白和SR蛋白竞争抑制外显子
无内含子的多外显子基因的互补链倾向于滞留于核内,这与它们特定的序列组成有关,其中富含嘌呤的剪切增强子可能会影响mRNA的出核过程。SR家族蛋白中也有部分可与富含GAA序列的剪切增强子motif结合,通过与mRNA出核因子NXF1相互作用参与mRNA出核。FLASH实验显示SAFB蛋白缺失后,SAFB靶向区域元件与SR蛋白的结合有所增加,受影响最大的则是正义L1转座子或其片段(图4a),说明SAFB蛋白是新生L1 RNP的重要组成部分,并且与SR蛋白存在竞争关系。亲和纯化质谱和免疫印迹等实验发现,存在一组hnRNP蛋白与SAFB蛋白相关,可以与SR蛋白动态竞争以抑制靶向RNA的外显子化。
SAFB蛋白与SR蛋白竞争结合富含嘌呤的motif,以防止L1、Tigger和假基因的同链插入对剪切过程造成干扰,而不干扰含有类似剪切增强子motif的外显子的剪切(图4a)。SAFB蛋白的抑制作用对转录本特征也有所偏好,长外显子(超过1kb)比平均大小的外显子(150nt左右)更容易受到抑制。在敲低SAFB处理细胞中,表达显著上调的外显子中,存在一类巨型蛋白编码盒式外显子,尽管表达它们存在组织特异性,但是序列都富含腺嘌呤及剪切增强子,这也反映了转座子L1和Tigger的序列组成偏好。
已有研究表明m6A修饰可能影响SAFB1与靶向RNA结合,并在长外显子以及自主转座子上呈现内源性富集。EMSA实验结果说明,相比于SAFB1,SR类蛋白更容易与RNA聚合酶II释放的未甲基化新生RNA结合;然而随着RNA在细胞核中逐渐甲基化,这种优势将向SAFB1转移(图4b)。因此,与平均大小的外显子相比,转座子L1 RNA和巨型蛋白编码盒式外显子更有可能通过m6A修饰受到SAFB的抑制调控。
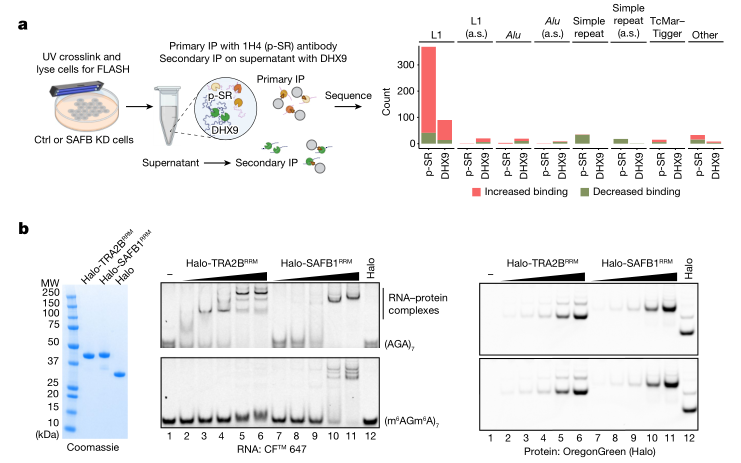
图4:m6A修饰介导SAFB蛋白和SR蛋白竞争抑制外显子
4. SAFB蛋白的调控功能解析
人类的SAFB靶向RNA有几个关键特征,包括含有一个长而连续的富含嘌呤的motif,以及SAFB蛋白缺失后内含子的外显子化。然而,转座子L1、Tiggle或巨型蛋白编码盒式外显子却表现不同。为了探究序列及功能是否保守,在小鼠及果蝇中进行验证发现,SAFB蛋白不仅抑制转座子L1,还同样影响LTR/ERV转座子家族,它们都具有相似的结构特征和表达模式。
SAFB蛋白在抑制转座子活性方面的作用机制具有深远的进化意义,并且可能源于调控嵌套基因表达、防止假基因外显子化以及调控巨型外显剪切子剪接的需要。转座子L1 ORF的腺嘌呤偏向性在脊椎动物中非常保守,这是自主转座子家族的序列共性,也意味着存在着一种迄今为止未知的进化压力来维持这一特性。
尽管SAFB蛋白在人体组织中广泛表达,但是在减数分裂后期的精子细胞中,SAFB1和SAFB2的表达量急剧下降(图5a)。通过对小鼠睾丸组织进行免疫荧光染色,证实了这一观察结果(图5c)。为了验证SAFB活性和转座子L1表达之间的反向关系,对仅在雄性种系中调节L1转录的Dnmt3c基因进行敲除,发现睾丸组织中ORF1p表达显著,但几乎仅限于SAFB1表达较低的分化细胞中(图5d)。这种种系特异性的关系为新的转座子插入提供了可能,因为它们可以遗传给下一代而不会给宿主造成突变负担。由于体细胞中的转座子插入致命却不可遗传,而Piwi相互作用RNA系统在生殖细胞系中处于活跃状态,因此将转座子与宿主间的冲突局限在精子发生过程中,对宿主和转座子来说是一种互利的策略。体细胞中被SAFB抑制的部分剪接事件在睾丸中被激活,与减数分裂后精子中SAFB的低表达一致,这一现象证实了前述机制(图5b)。这使人联想到了抵抗外部病原体时先天免疫和适应性免疫的分工,由此说明SAFB蛋白作为基于RNA、模式引导的非获得性防御系统,能够抑制转座子,并与种系中基于RNA的适应性Piwi互作RNA通路形成互补。
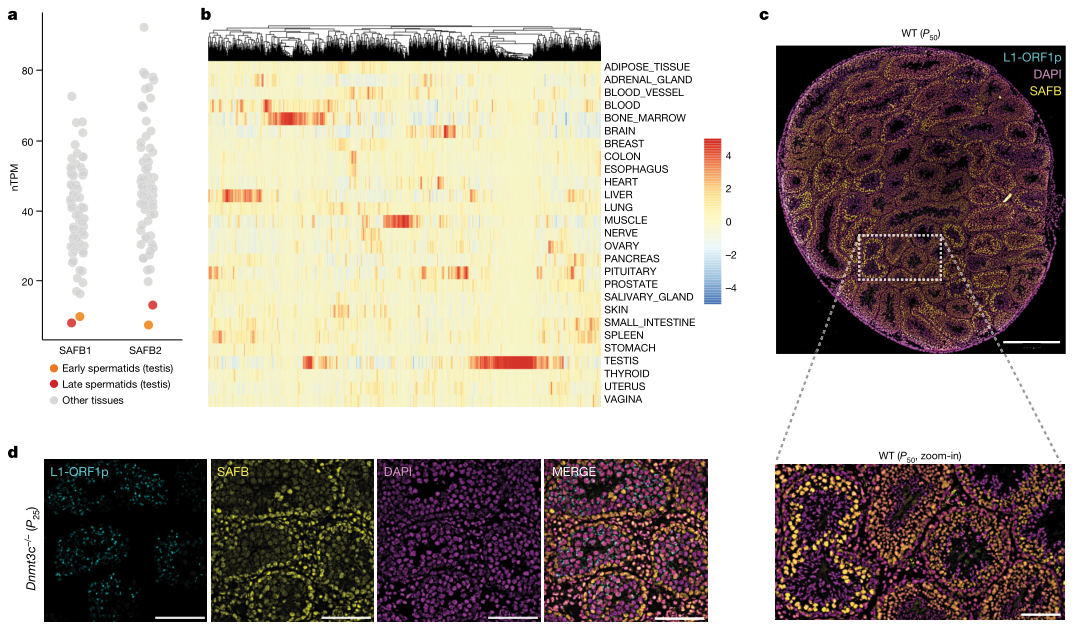
图5:SAFB蛋白的调控功能解析
研究结论
本文通过Direct RNA测序及多种生理生化方法,研究了SAFB蛋白对转座子活性的调控作用及其在进化中的意义,发现SAFB蛋白在调控转座子活性和维护基因组稳定性方面扮演着关键角色,揭示了其内在作用机制与进化压力和环境响应密切相关。
参考文献:
Ilık İ A, Glažar P, Tse K, et al. Autonomous transposons tune their sequences to ensure somatic suppression[J]. Nature, 2024: 1-9.


