 027-62435310 |
027-62435310 |
 service@speedracings.com |
service@speedracings.com |
高分文献解读 | Nature Plants期刊发表铁线蕨基于基因组组装揭示真叶植物的起源和茉莉素进化机制
AG代理基因长期致力于基因组测序和组装技术的研发,实现了N50>100Kb超长序列的建库测序稳定产出并开发了相应T2T组装全流程,已完成多类型物种的T2T基因组组装,有丰富项目经验,期待为您提供从ONT超长测序到物种完整T2T基因组组装的全流程服务。
详情请发送邮件咨询:service@speedracings.com
研究背景
真叶植物涵盖了几乎所有现存的植物,包括蕨类植物和种子植物。蕨类植物是地球陆生植物中最古老和最具生态意义的维管植物谱系,其系统发育位置与种子植物最为接近,故解码蕨类植物的基因组是深入了解真叶植物起源和种子植物进化的关键。本研究中,作者组装得到了蕨类植物模型铁线蕨的染色体水平基因组。基因共表达网络分析发现,蕨类植物同型孢子发育与种子植物的花粉具有较高的遗传相似性。此外,该研究还分析了蕨类植物的防御反应,扩展了对植物内源性生物活性茉莉酸盐进化和多样性的理解。最后,作者还将蕨类植物的基因组与其他陆生植物的基因组进行了比较,揭示了基因家族的变化,这些基因家族对于真叶植物分支内的进化新奇性非常重要。总之,这些结果为蕨类植物基因组的进化和功能研究以及真叶植物的起源和进化研究奠定了基础。

文章题目:The genome of homosporous maidenhair fern sheds light on the euphyllophyte evolution and defences
发表期刊:Nature Plants(IF=17.352)
发表时间:2022.09.01
主要研究结果
1.铁线蕨基因组组装和注释
为了获得高质量的蕨类植物基因组,该研究首先构建了一个完全纯和的二倍体铁线蕨。然后,基于K-mer分析(4.95Gb)、PacBio测序数据(126X)、illumina双端测序数据(68X)、Hi-C数据(551.74Mb有效数据)以及BioNano光学图谱数据(141.32X),最终组装获得了4.83Gb的铁线蕨染色体水平基因组,覆盖率97.58%,Contig N50达16.22Mb,且被挂载到了30条染色体上,挂载率高达98.79%(图1)。进一步评估基因组质量得:二代比对率95.44%,97.90%的表达序列标签可以映射到染色体序列上,此外,BioNano比对率97.05%且仅有2个嵌合contig、没有大于5Kb的indel。这些结果均表明,本研究组装得到的基因组是高度连续和完整的。
对该基因组注释发现,铁线蕨基因组高度重复,注释到的重复序列占基因组长度的85.25%,且这些重复元件在不同的染色体上的分布是不同的。注释还发现,rRNA大多数嵌入在富含Gypsy和Copia的重复区域内,最显著的rRNA簇出现在3号和23号染色体上。此外,作者通过基因结构预测,高度可信地注释到了31244个核编码基因。
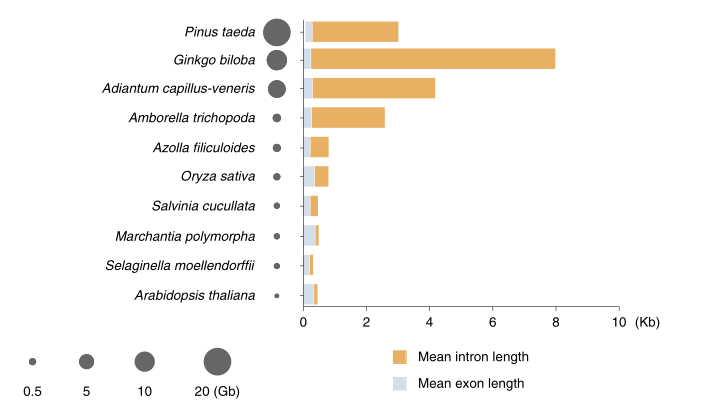
图1 铁线蕨基因组从头组装
2.铁线蕨缺乏最近的全基因组复制事件
植物基因组大小扩增的主要原因之一是全基因组复制事件的发生,为了研究铁线蕨是否发生了WGD事件,作者进行了基因组内共线性分析,结果显示,7个共线性区块中仅有96个共线性旁系同源基因,仅占总基因的0.31%,说明共线性块的分布十分稀疏。此外,KS分布图也没有显示出全基因组复制事件对应的峰值。故说明铁线蕨基因组中最近没有发生WGD事件,即WGD不是其基因组大小扩增的主要原因。然而,旁系同源基因组的KS分布显示了一个KS>2的模糊峰值,为了探究其是否代表在PTERa之前发生了古WGD事件,作者又采用了一种叫做ksrates的新方法来进行KS分析,结果,校正后的数据支持了WGD事件的存在,但仅代表所有核心薄囊物种均存在的古WGD事件,而没有在其基因组中出现新一轮的WGD。
3.重复元件是区分同孢蕨类和异孢蕨类的基因组特征
作者发现,铁线蕨的重复元件基因组占比(85.25%)远大于其他两种异孢蕨类(A. filiculoides-41.60% and S. cucullate-34.62%),其主要成分LTR-RTs也是如此。因此,作者计算了核苷酸距离D来推断插入和消除率。结果发现,铁线蕨早期存在爆发式以及持续性的LTR-RT插入,同时其也有较高的消除率,但总的来说,持续性的插入仍然导致铁线蕨LTR-RT的显著累积。所以,这些发现表明重复元件的扩张是同孢蕨类植物基因组扩张的基础。同时,该研究发现铁线蕨的平均基因组大小远大于异孢蕨类的平均基因组大小,但平均外显子大小和每个基因的平均外显子数量相似,因此,作者认为铁线蕨基因结构的显著特征是内含子大小的增大(图2),对内含子中重复元件的分析还发现,LTR-RT和LINE是导致铁线蕨内含子长度增大的重要重复元件,以往的研究表明,这种情况在其他较大的植物基因组如银杏中也有观察到。这些结果都表明,在这些植物中,基因组大小可能与平均内含子的长度呈正相关。
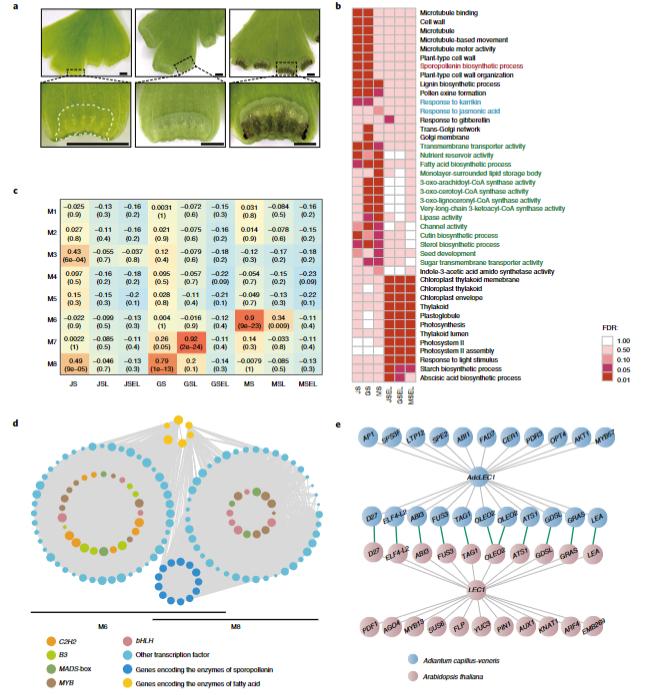
图2 10种陆地植物的内含子和外显子长度分布
4.同型孢子发育调控机制
结合转录组分析,研究人员进一步揭示了同型孢子囊从初生期到成熟期的调控机制,首次发现同型孢子发育与异性孢子的花粉具有相似性。通过对同型孢子共表达模块中节点基因的挖掘和比较网络分析,发现LEC1基因在成熟阶段调控同化物积累,而其种子植物同源基因也有相同的功能,是种子发育的核心调控因子,证明了LEC1调控网络在种子起源前至少300万年已经形成,且具有保守的同化物积累调控功能(图3)。在蕨类植物中,该网络在孢子发育时期发挥作用,而在种子植物中,则被招募到种子发育过程中,赋予了种子同化物积累这一重要的生理性状。这些结果为我们更好地理解种子起源提供了新的见解。
具体来说,先前的研究表明,LEC1可以招募ABI3和FUS3形成基因调节网络,以执行适应性生理特征,如贮藏化合物积累、脱水耐性和休眠获得,并作为种子植物种子发育的主调节器。在铁线蕨中,同源基因AdcLEC1在成熟的同孢子囊中表现出明确的特异性表达模式,并且在成熟的与同孢子囊相关的M6模块中与AdcABI3和AdcFUS3共同表达;这与拟南芥种子中ABI3和FUS3上的LEC1激活相似,这意味着AdcLEC1基因调控网络可能在同孢子囊发育中发挥作用。为了进一步评估同孢子体和种子中LEC1调节网络的功能保守性,作者筛选出了1390个与AdcLEC1共表达的基因,随后通过BLAST将它们与拟南芥中的1515个AtLEC1共表达基因进行比较,结果发现,在拟南芥和铁线蕨LEC1网络中检测到一些编码种子特异性储存蛋白的同源物,如AdcOLEOSIN和AdcLEA,这说明了LEC1介导的储存蛋白积累具有类似的分子机制。如图3e所示,LEC1相关网络的核心成分在在铁线蕨和拟南芥中都是保守的。此外,以往的其他研究也暗示了LEC1在同化积累上的功能保守性。
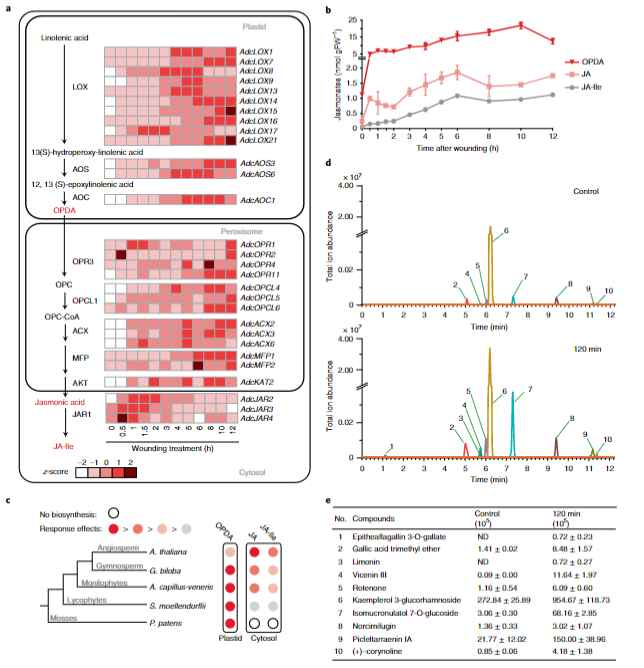
图3 铁线蕨同型孢子发育过程中的表达谱和基因网络分析
5.茉莉酸信号在植物防御中的分子进化
蕨类植物对虫害和病原体具有很高的抵抗力,其虫害发生率比开花植物低30倍左右。对蕨类植物的研究表明,茉莉酸信号可能是蕨类植物强大的病原菌抗性的重要因素。为了深入分析蕨类植物中的JA信号,研究人员对蕨类植物的叶子进行了机械性伤害以模仿昆虫的食草性。转录组分析表明,大多数JA生物合成基因都被诱导表达且持续上升,这表明伤害可以出发蕨类植物快速的茉莉酸生物合成且这种创伤诱导是持续性的。以往的研究表明,JA-lle是种子植物中的主要活性分子,而石松类和苔藓类植物中则主要以OPDA的形式出现。但该研究转录组分析表明,大多数JA生物合成基因都被诱导表达且持续上升,这表明伤害可以出发蕨类植物快速的茉莉酸生物合成且这种创伤诱导是持续性的(图4)。
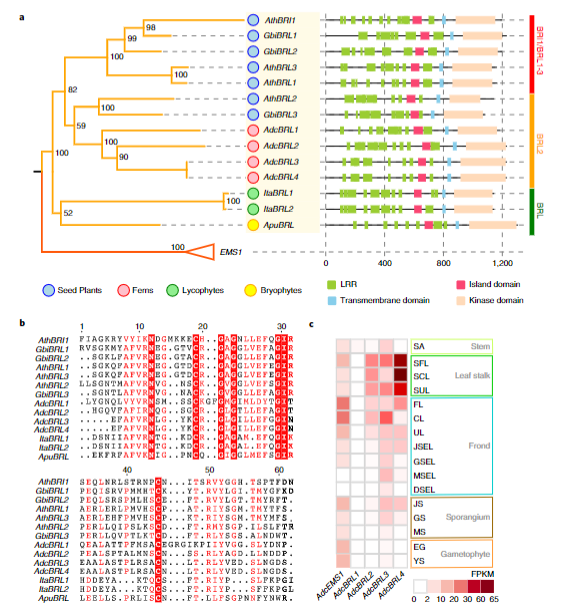
图4 铁线蕨JA合成与信号转导通路解析
6.BRI1-BRL基因家族在真叶植物中的扩张与真叶的起源演化
通过对真叶植物各进化节点的祖先基因推断,首次发现真叶植物的最近共同祖先中,BRI1-BRL基因家族出现了明显扩张。研究显示了BRI1-BRL基因家族在真叶植物中发生过两次复制,从而形成了真叶植物共有的BRL2亚家族,以及种子植物中特异扩张的BRL1/BRL1-3亚家族(图5)。铁线蕨BRL2亚家族成员的组织表达分析也验证了这些基因在维管束分布丰富的叶柄和叶片中表达显著。这些结果显示了BRI1-BRL基因家族的扩张很可能与真叶维管系统的起源有关,是推动真叶植物维管系统复杂化进而产生真叶的分子动力。
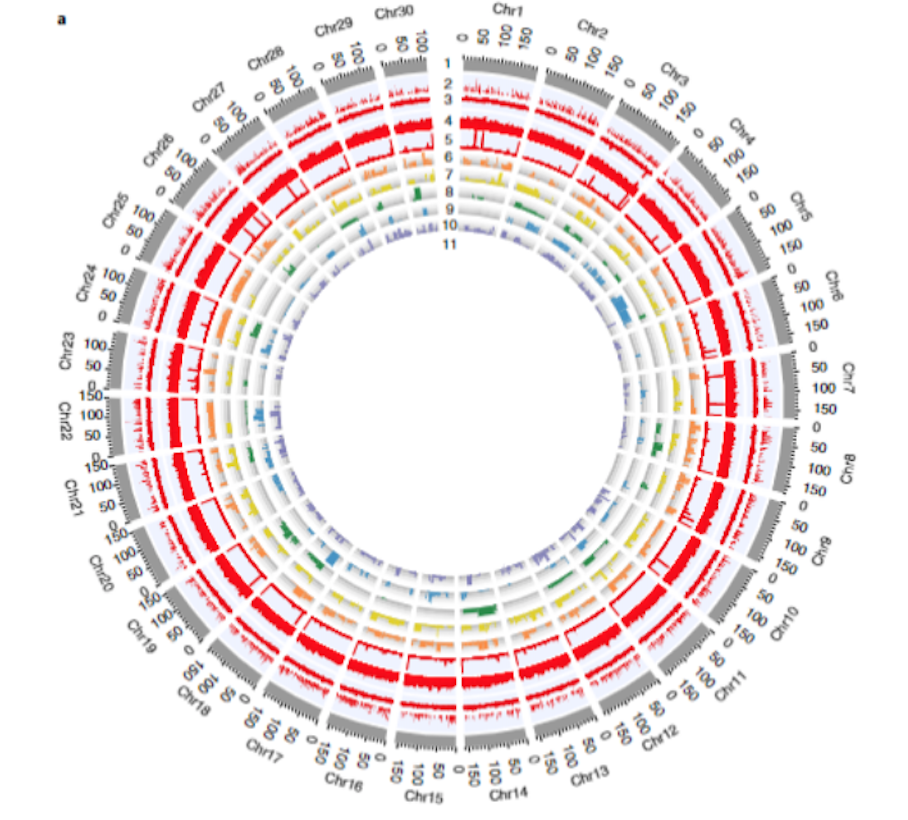
图5 BRI1-BRL基因家族进化
研究结论:
系统发育位置和其同孢性质使铁线蕨基因组成为一个有价值的参考,有助于揭示关键的进化事件,包括同孢囊发育和突出的病原体抗性。该研究根据同孢囊的遗传基础进行推断,揭示了同型孢子发育过程与异型孢子花粉的相似性。作为代表同型孢子蕨类的模式物种,铁线蕨的高质量基因组为研究现存蕨类的起源以及基因家族进化等相关的一系列特征提供了有意义的模型。


